
Adlay Seed (Coix lacryma-jobi L. var. Ma-yuen Stapf.) Ethanolic Extract Fractions and Subfractions Induce Cell Cycle Arrest and Apoptosis in Human Breast and Cervical Cancer Cell Lines

 , ,
, ,  , , , and
, , , and 
Abstract
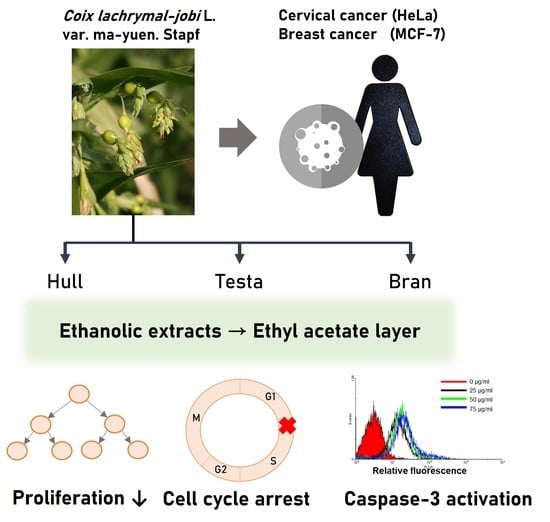
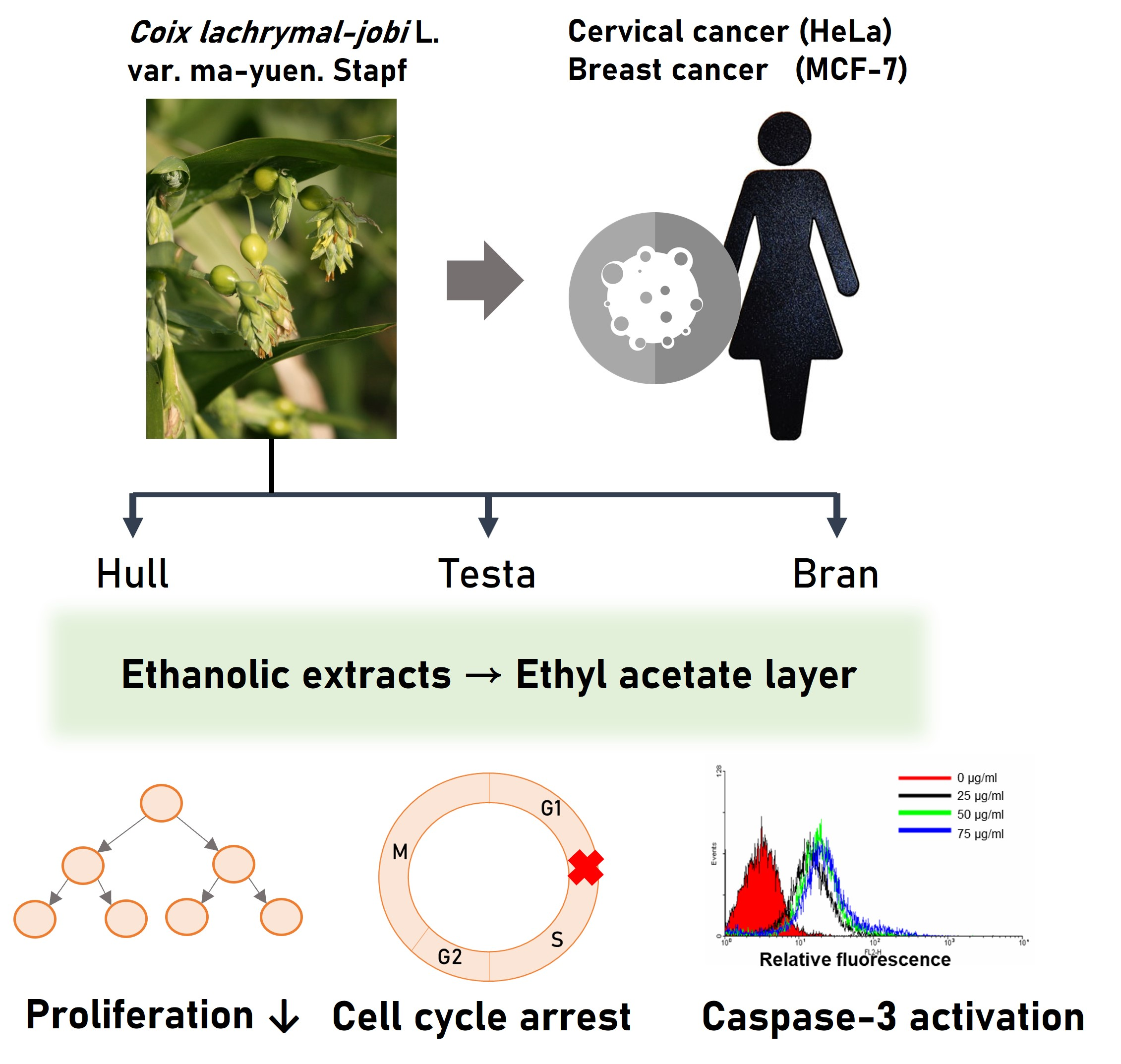
:
1. Introduction
2. Results
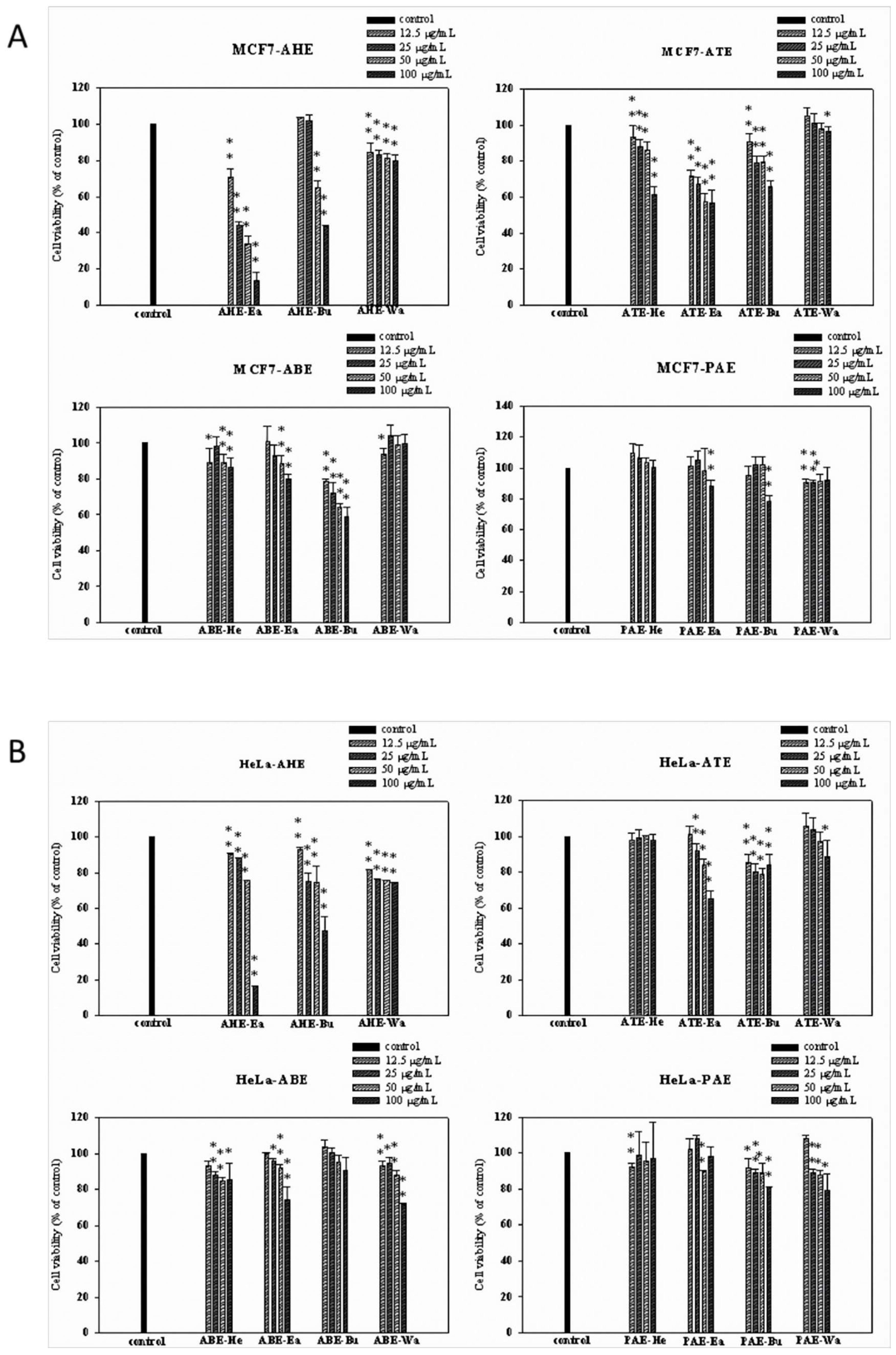
2.1. Effects of Different Parts of the Adlay Seed and Its Ethanolic Extract Fractions on Cancer Cell Viability
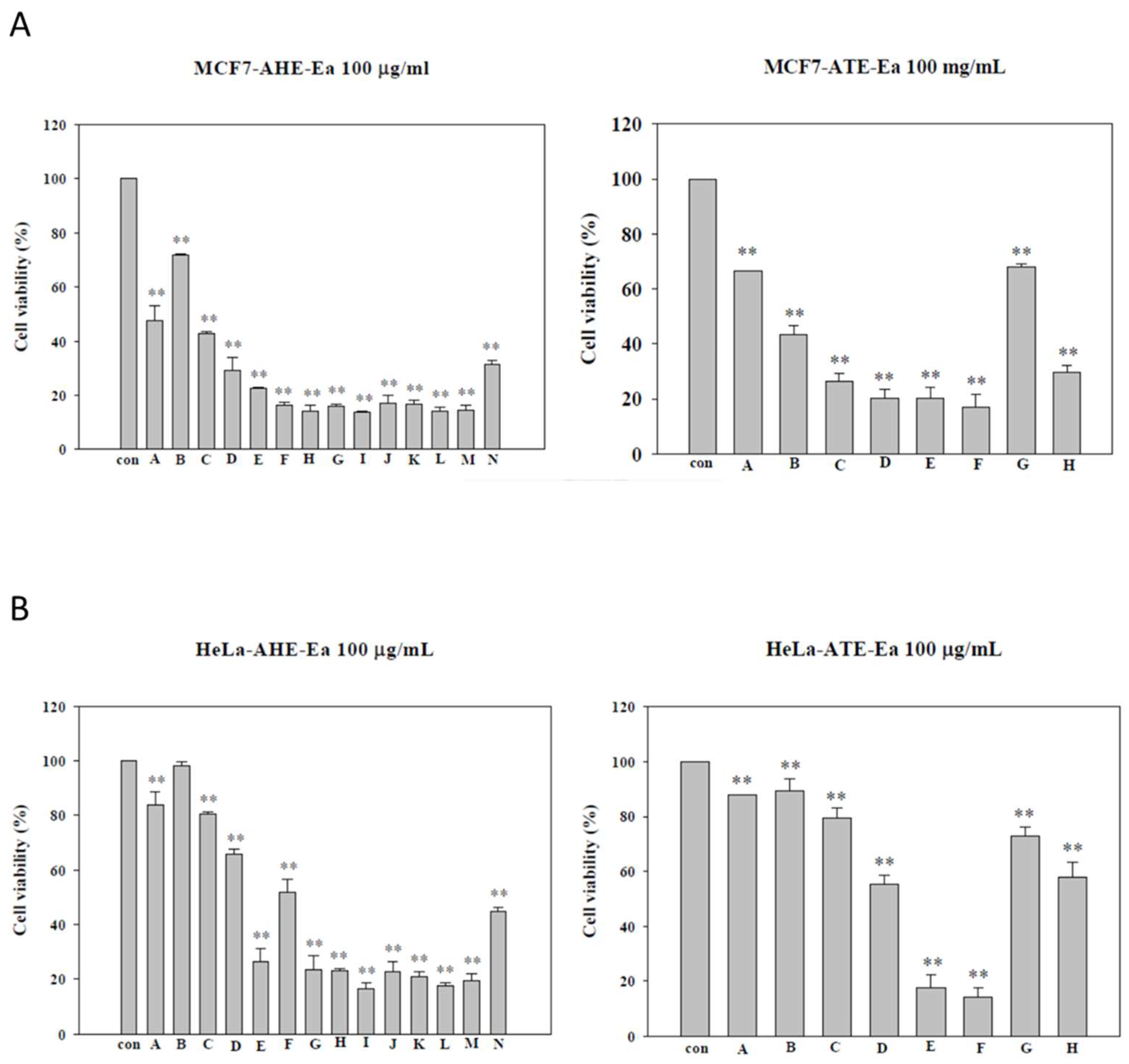
2.2. Effects of Different Subfractions of Adlay Hull and Testa Extracts on Cancer Cell Viability
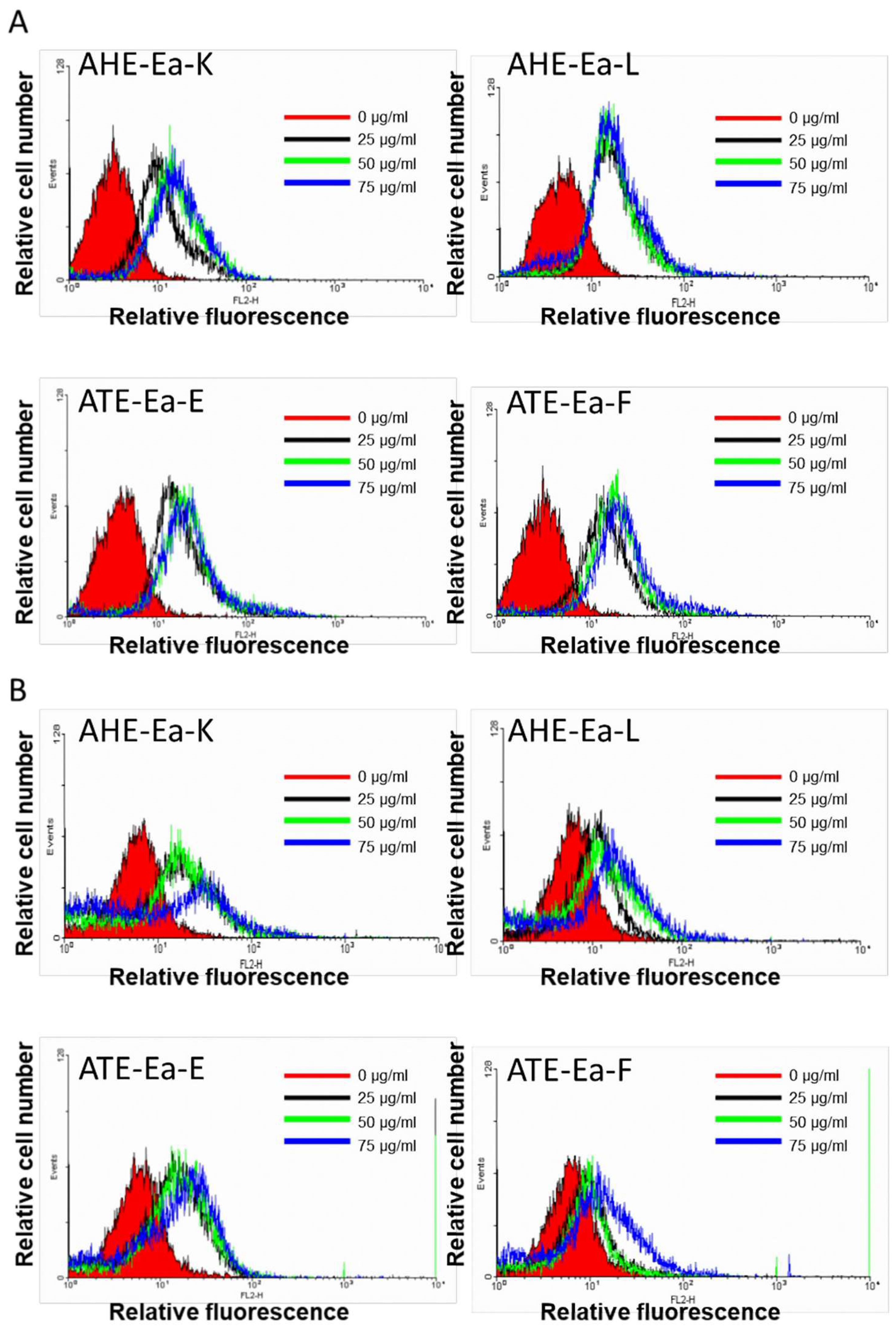
2.3. Effects of AHE-Ea-K, AHE-Ea-L, ATE-Ea-E and ATE-Ea-F Subfractions on Cancer Cell Cycle Distribution
2.4. Effects of AHE-Ea-K, AHE-Ea-K L, ATE-Ea-E and ATE-Ea-F on Caspase-3 Activity
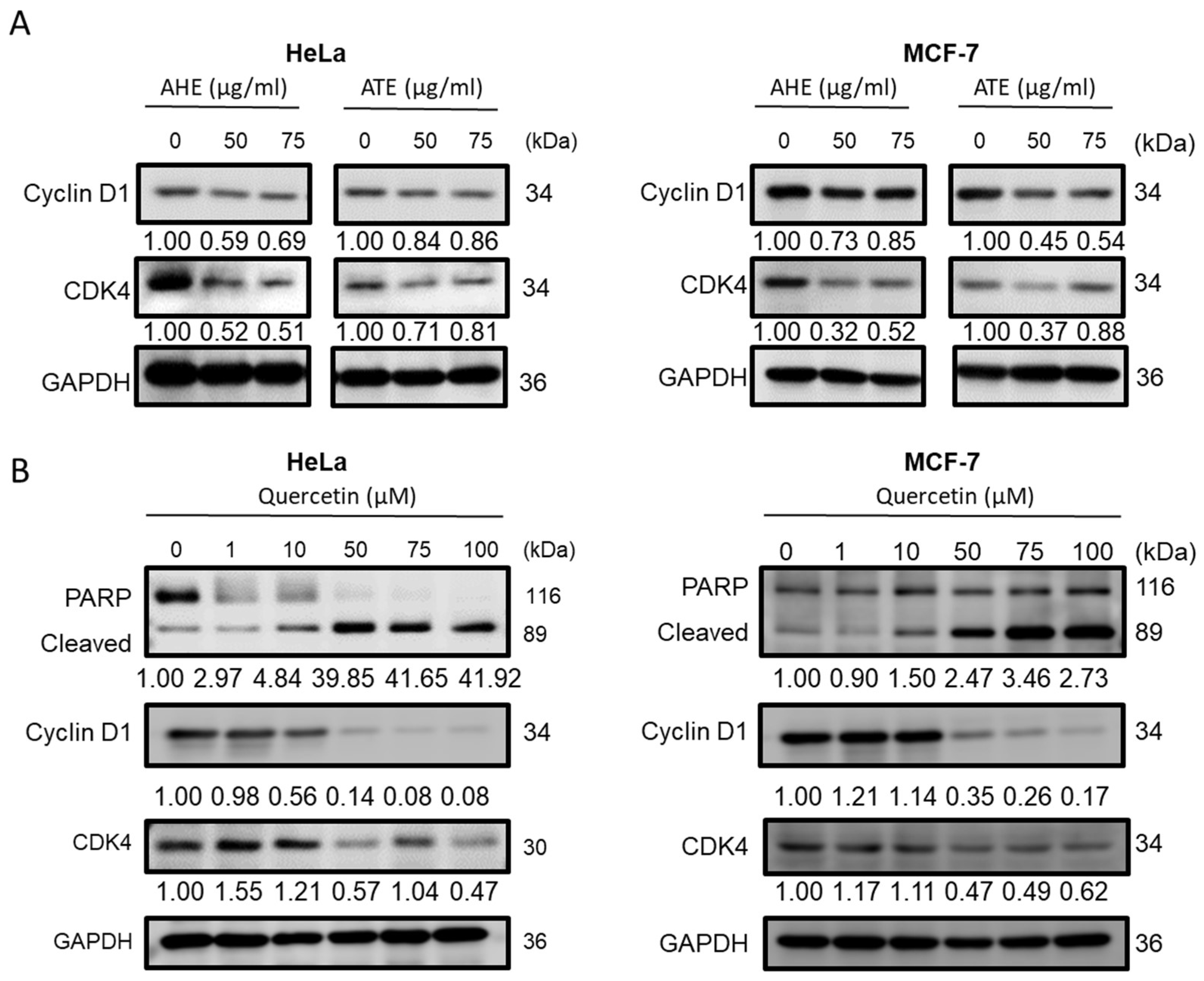
2.5. Anticancer Properties of Adlay Extract and its Active Components
3. Discussion
4. Materials and Methods
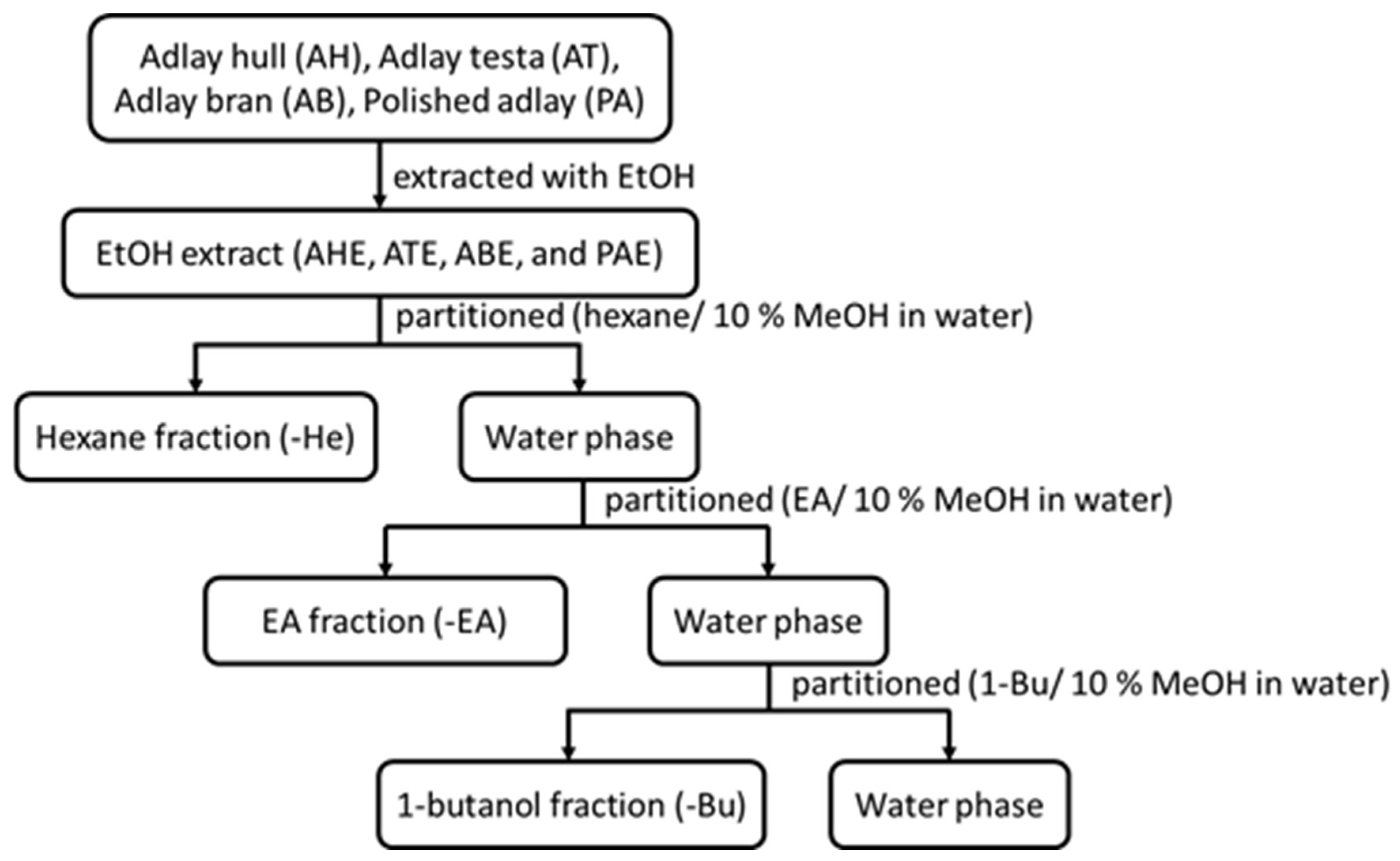
4.1. Adlay Extract Preparation
4.2. Cell Line and Culture Conditions
4.3. MTT Assay
4.4. Cell Cycle Distribution
4.5. Caspase-3 Activity Analysis
4.6. Protein Extraction and Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sultova, E.; Westphalen, C.B.; Jung, A.; Kumbrink, J.; Kirchner, T.; Mayr, D.; Rudelius, M.; Ormanns, S.; Heinemann, V.; Metzeler, K.H.; et al. NGS-guided precision oncology in metastatic breast and gynecological cancer: First experiences at the CCC Munich LMU. Arch. Gynecol. Obstet. 2021, 303, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Shen, R.; Richard, S.; Liu, Y.; Jalalirad, M.; Cleary, M.P.; D’Assoro, A.B.; Gradilone, S.A.; Yang, D.Q. Inhibition of triple-negative breast cancer proliferation and motility by reactivating p53 and inhibiting overactivated Akt. Oncol. Rep. 2022, 47, 41. [Google Scholar] [CrossRef]
- Fishelson, Z.; Attali, G.; Mevorach, D. Complement and apoptosis. Mol. Immunol. 2001, 38, 207–219. [Google Scholar] [CrossRef]
- Lin, P.H.; Chiang, Y.F.; Shieh, T.M.; Chen, H.Y.; Shih, C.K.; Wang, T.H.; Wang, K.L.; Huang, T.C.; Hong, Y.H.; Li, S.C.; et al. Dietary Compound Isoliquiritigenin, an Antioxidant from Licorice, Suppresses Triple-Negative Breast Tumor Growth via Apoptotic Death Program Activation in Cell and Xenograft Animal Models. Antioxidants 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, D.W.; Ali, A.; Thornberry, N.A.; Vaillancourt, J.P.; Ding, C.K.; Gallant, M.; Gareau, Y.; Griffin, P.R.; Labelle, M.; Lazebnik, Y.A.; et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 1995, 376, 37–43. [Google Scholar] [CrossRef]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef]
- Huang, Y.J.; Chang, C.C.; Wang, Y.Y.; Chiang, W.C.; Shih, Y.H.; Shieh, T.M.; Wang, K.L.; Ali, M.; Hsia, S.M. Adlay Testa (Coix lachryma-jobi L. var. Ma-yuen Stapf.) Ethanolic Extract and Its Active Components Exert Anti-Proliferative Effects on Endometrial Cancer Cells via Cell Cycle Arrest. Molecules 2021, 26, 1966. [Google Scholar] [CrossRef]
- Chung, C.P.; Lee, M.Y.; Hsia, S.M.; Chiang, W.; Kuo, Y.H.; Hsu, H.Y.; Lin, Y.L. Suppression on allergic airway inflammation of dehulled adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) in mice and anti-degranulation phytosterols from adlay bran. Food Funct. 2021, 12, 12788–12799. [Google Scholar] [CrossRef]
- Huang, C.C.; Lin, T.C.; Liu, C.H.; Hu, H.C.; Yu, S.Y.; Wu, S.J.; Yen, M.H.; Tsai, Y.H.; Chang, F.R. Lipid Metabolism and its Mechanism Triggered by Supercritical CO(2) Extract of Adlay (Coix lacryma-jobi var. ma-yuen (Rom. Caill.) Stapf) Bran in High-Fat Diet Induced Hyperlipidemic Hamsters. Front. Pharmacol. 2021, 12, 785944. [Google Scholar] [CrossRef]
- Yeh, P.H.; Chiang, W.; Chiang, M.T. Effects of dehulled adlay on plasma glucose and lipid concentrations in streptozotocin-induced diabetic rats fed a diet enriched in cholesterol. Int. J. Vitam. Nutr. Res. 2006, 76, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Chen, Y.C.; Chen, H.Y.; Chiang, Y.F.; Ali, M.; Chiang, W.; Chung, C.P.; Hsia, S.M. Ethanolic Extracts of Adlay Testa and Hull and Their Active Biomolecules Exert Relaxing Effect on Uterine Muscle Contraction through Blocking Extracellular Calcium Influx in Ex Vivo and In Vivo Studies. Biomolecules 2021, 11, 887. [Google Scholar] [CrossRef]
- Hsia, S.M.; Yeh, C.L.; Kuo, Y.H.; Wang, P.S.; Chiang, W. Effects of adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) hull extracts on the secretion of progesterone and estradiol in vivo and in vitro. Exp. Biol. Med. (Maywood) 2007, 232, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.C.; Shih, M.C.; Kuo, Y.H.; Chiang, W. Antagonism of free-radical-induced damage of adlay seed and its antiproliferative effect in human histolytic lymphoma U937 monocytic cells. J. Agric. Food Chem. 2001, 49, 1564–1570. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Huang, Y.C.; Hung, W.C. Antiproliferative and chemopreventive effects of adlay seed on lung cancer in vitro and in vivo. J. Agric. Food Chem. 2003, 51, 3656–3660. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Shih, C.K.; Yen, Y.T.; Chiang, W.; Hsia, S.M. Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf.) Hull Extract and Active Compounds Inhibit Proliferation of Primary Human Leiomyoma Cells and Protect against Sexual Hormone-Induced Mice Smooth Muscle Hyperproliferation. Molecules 2019, 24, 1556. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Huang, L.H.; Chiang, W.; Hsia, S.M. Hexane fraction of adlay (Coix lachryma-jobi L.) testa ethanolic extract inhibits human uterine sarcoma cancer cells growth and chemosensitizes human uterine sarcoma cells to doxorubicin. Phytomedicine 2018, 47, 69–80. [Google Scholar] [CrossRef]
- Chiu, L.C.; Ho, T.S.; Wong, E.Y.; Ooi, V.E. Ethyl acetate extract of Patrinia scabiosaefolia downregulates anti-apoptotic Bcl-2/Bcl-X(L) expression, and induces apoptosis in human breast carcinoma MCF-7 cells independent of caspase-9 activation. J. Ethnopharmacol. 2006, 105, 263–268. [Google Scholar] [CrossRef]
- Fang, N.; Li, Q.; Yu, S.; Zhang, J.; He, L.; Ronis, M.J.; Badger, T.M. Inhibition of growth and induction of apoptosis in human cancer cell lines by an ethyl acetate fraction from shiitake mushrooms. J. Altern. Complement. Med. 2006, 12, 125–132. [Google Scholar] [CrossRef] [Green Version]
- van Meeuwen, J.A.; Korthagen, N.; de Jong, P.C.; Piersma, A.H.; van den Berg, M. (Anti)estrogenic effects of phytochemicals on human primary mammary fibroblasts, MCF-7 cells and their co-culture. Toxicol. Appl. Pharmacol. 2007, 221, 372–383. [Google Scholar] [CrossRef]
- Xia, W.B.; Xue, Z.; Li, S.; Wang, S.J.; Yang, Y.C.; He, D.X.; Ran, G.L.; Kong, L.Z.; Shi, J.G. Chemical constituents from tuber of Cremastra appendiculata. Zhongguo Zhong Yao Za Zhi 2005, 30, 1827–1830. [Google Scholar] [PubMed]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Huang, T.-C.; Shieh, T.-M.; Wu, C.-H.; Lin, L.-C.; Hsia, S.-M. Isoliquiritigenin Induces Autophagy and Inhibits Ovarian Cancer Cell Growth. Int. J. Mol. Sci. 2017, 18, 2025. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AHE-Ea | MCF-7 (μg/mL) | HeLa (μg/mL) |
|---|---|---|
| A | 100.5 | >200 |
| B | 146.3 | >200 |
| C | 81.9 | >200 |
| D | 43.8 | 129.2 |
| E | 34.8 | 38.7 |
| F | 31.4 | 108.3 |
| G | 14.2 | 49.63 |
| H | 14.2 | 47.7 |
| I | 14.4 | 34.7 |
| J | 32.9 | 38.2 |
| K | 13.7 | 32.4 |
| L | 13.7 | 38.4 |
| M | 21.2 | 98.8 |
| N | 60.9 | 1.8 |
| ATE-Ea | MCF-7 (μg/mL) | HeLa (μg/mL) |
|---|---|---|
| A | 121.7 | 151.9 |
| B | 75.7 | 145.4 |
| C | 46.4 | 133.3 |
| D | 54.0 | 114.8 |
| E | 25.0 | 58.2 |
| F | 16.3 | 34.1 |
| G | 130.5 | 170.9 |
| H | 139.6 | 110.4 |
| MCF-7 | HeLa | |||||
|---|---|---|---|---|---|---|
| AHE-Ea-K (μg/mL) | G0/G1 | S | G2/M | G0/G1 | S | G2/M |
| 0 | 46.9 ± 0.6 b | 26.8 ± 0.1 a | 26.4 ± 0.7 a | 42.9 ±1.5 a | 34.0 ± 2.4 d | 23.2 ± 10 a |
| 25 | 62.8 ± 0.1 ab | 16.8 ± 0.2 b | 20.4 ± 0.1 a | 42.7 ± 3.6 a | 44.3 ± 1.5 c | 13.0 ± 1.6 ab |
| 50 | 69.0 ± 1.0 a | 9.9 ± 0.3 d | 21.1 ± 1.3 a | 30.2 ± 1.5 b | 50.1 ± 3.4 b | 19.7 ± 4.9 ab |
| 75 | 72.4 ± 3.2 a | 13.4 ± 0.2 c | 14.2 ± 3.0 a | 36.0 ± 5.5 ab | 61.8 ± 6.9 a | 2.2 ± 1.8 b |
| AHE-Ea-L (μg/mL) | G0/G1 | S | G2/M | G0/G1 | S | G2/M |
| 0 | 47.3 ± 0.7 c | 32.2 ± 2.0 a | 20.4 ± 1.2 a | 56.2 ± 5.5 a | 24.7 ± 5.8 c | 19.2 ± 2.9 a |
| 25 | 60.0 ± 2.0 b | 18.8 ± 1.5 b | 21.2 ± 0.7 a | 59.7 ± 8.7 a | 30.4 ± 3.8 c | 9.9 ± 8.3 b |
| 50 | 63.7 ± 1.7 b | 14.9 ± 0.2 b | 21.4 ± 2.0 a | 38.7 ± 1.7 b | 41.1 ± 0.5 b | 20.2 ± 2.1 a |
| 75 | 76.1 ± 4.3 a | 4.0 ± 2.5 c | 19.9 ± 6.7 a | 39.4 ± 2.5 b | 59.3 ± 1.6 a | 1.3 ± 0.9 c |
| ATE-Ea-E (μg/mL) | G0/G1 | S | G2/M | G0/G1 | S | G2/M |
| 0 | 43.4 ± 2.9 c | 45.9 ± 1.2 a | 10.7 ± 1.8 a | 35.2 ± 6.3 c | 53.7 ± 8.0 a | 11.1 ± 3.9 a |
| 25 | 49.7 ± 8.6 b | 40.0 ± 9.3 b | 10.4 ± 4.7 a | 40.2 ± 7.1 b | 47.6 ± 5.6 ab | 12.2 ± 1.6 a |
| 50 | 56.1 ± 0.6 a | 42.8 ± 0.6 a | 1.1 ± 1.2 b | 47.7 ± 1.9 a | 44.6 ± 0.9 bc | 7.7 ± 2.8 a |
| 75 | 58.3 ± 1.4 a | 39.1 ± 0.6 a | 2.6 ± 0.8 b | 47.7 ± 1.2 a | 39.3 ± 0.8 c | 13.0 ± 2.0 a |
| ATE-Ea-F (μg/mL) | G0/G1 | S | G2/M | G0/G1 | S | G2/M |
| 0 | 51.6 ± 5.0 c | 36.5 ± 5.9 a | 11.9 ± 3.0 a | 42.1 ± 9.6 c | 42.4 ± 8.5 a | 15.3 ± 3.0 c |
| 25 | 51.4 ± 3.8 b | 37.1 ± 6.3 a | 11.5 ± 2.9 a | 50.5 ± 1.6 b | 36.2 ± 2.5 a | 13.3 ± 1.0 c |
| 50 | 65.3 ± 6.6 b | 24.1 ± 5.1 b | 10.6 ± 1.9 a | 54.1 ± 3.0 ab | 21.8 ± 3.7 b | 24.1 ± 6.4 a |
| 75 | 85.7 ± 0.6 a | 4.9 ± 2.1 c | 9.4 ± 1.5 a | 59.3 ± 2.7 a | 21.0 ± 2.1 b | 19.7 ± 0.6 b |
| MCF-7 | HeLa | |
|---|---|---|
| AHE-Ea-K (μg/mL) | Caspase-3 Positive Cells (%) | Caspase-3 Positive Cells (%) |
| 0 | 0.55 ± 0.05 d | 2.82 ± 0.18 d |
| 25 | 51.57 ± 0.38 c | 29.37 ± 2.10 c |
| 50 | 53.96 ± 0.01 b | 33.81 ± 0.75 b |
| 75 | 57.83 ± 0.25 a | 37.94 ± 0.11 a |
| AHE-Ea-L (μg/mL) | Caspase-3 Positive Cells (%) | Caspase-3 Positive Cells (%) |
| 0 | 0.55 ± 0.05 d | 2.82 ± 0.18 d |
| 25 | 36.37 ± 0.18 c | 7.10 ± 0.01 c |
| 50 | 38.94 ± 0.57 b | 10.28 ± 0.47 b |
| 75 | 39.67 ± 0.32 a | 34.80 ± 0.86 a |
| AHE-Ea-E (μg/mL) | Caspase-3 Positive Cells (%) | Caspase-3 Positive Cells (%) |
| 0 | 0.56 ± 0.17 d | 2.82 ± 0.18 d |
| 25 | 17.95 ± 0.81 c | 29.06 ± 0.23 c |
| 50 | 34.04 ± 0.54 b | 34.75 ± 0.15 b |
| 75 | 37.35 ± 0.16 a | 38.76 ± 0.62 a |
| AHE-Ea-F (μg/mL) | Caspase-3 Positive Cells (%) | Caspase-3 Positive Cells (%) |
| 0 | 0.56 ± 0.17 d | 2.82 ± 0.18 d |
| 25 | 27.43 ± 0.19 c | 7.68 ± 0.23 c |
| 50 | 36.99 ± 0.13 b | 7.92 ± 0.15 b |
| 75 | 53.42 ± 0.35 a | 25.85 ± 0.62 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, Y.-F.; Chung, C.-P.; Lin, J.-H.; Chiang, W.; Chen, H.-Y.; Ali, M.; Shih, Y.-H.; Wang, K.-L.; Huang, T.-C.; Chang, H.-Y.; et al. Adlay Seed (Coix lacryma-jobi L. var. Ma-yuen Stapf.) Ethanolic Extract Fractions and Subfractions Induce Cell Cycle Arrest and Apoptosis in Human Breast and Cervical Cancer Cell Lines. Molecules 2022, 27, 3984. https://doi.org/10.3390/molecules27133984
Chiang Y-F, Chung C-P, Lin J-H, Chiang W, Chen H-Y, Ali M, Shih Y-H, Wang K-L, Huang T-C, Chang H-Y, et al. Adlay Seed (Coix lacryma-jobi L. var. Ma-yuen Stapf.) Ethanolic Extract Fractions and Subfractions Induce Cell Cycle Arrest and Apoptosis in Human Breast and Cervical Cancer Cell Lines. Molecules. 2022; 27(13):3984. https://doi.org/10.3390/molecules27133984
Chicago/Turabian StyleChiang, Yi-Fen, Cheng-Pei Chung, Jing-Hui Lin, Wenchang Chiang, Hsin-Yuan Chen, Mohamed Ali, Yin-Hwa Shih, Kai-Lee Wang, Tsui-Chin Huang, Hsin-Yi Chang, and et al. 2022. "Adlay Seed (Coix lacryma-jobi L. var. Ma-yuen Stapf.) Ethanolic Extract Fractions and Subfractions Induce Cell Cycle Arrest and Apoptosis in Human Breast and Cervical Cancer Cell Lines" Molecules 27, no. 13: 3984. https://doi.org/10.3390/molecules27133984