
Structural Characterization and In-Vitro Antioxidant and Immunomodulatory Activities of Polysaccharide Fractions Isolated from Artemisia annua L.


Abstract
:1. Introduction
2. Results and Discussion
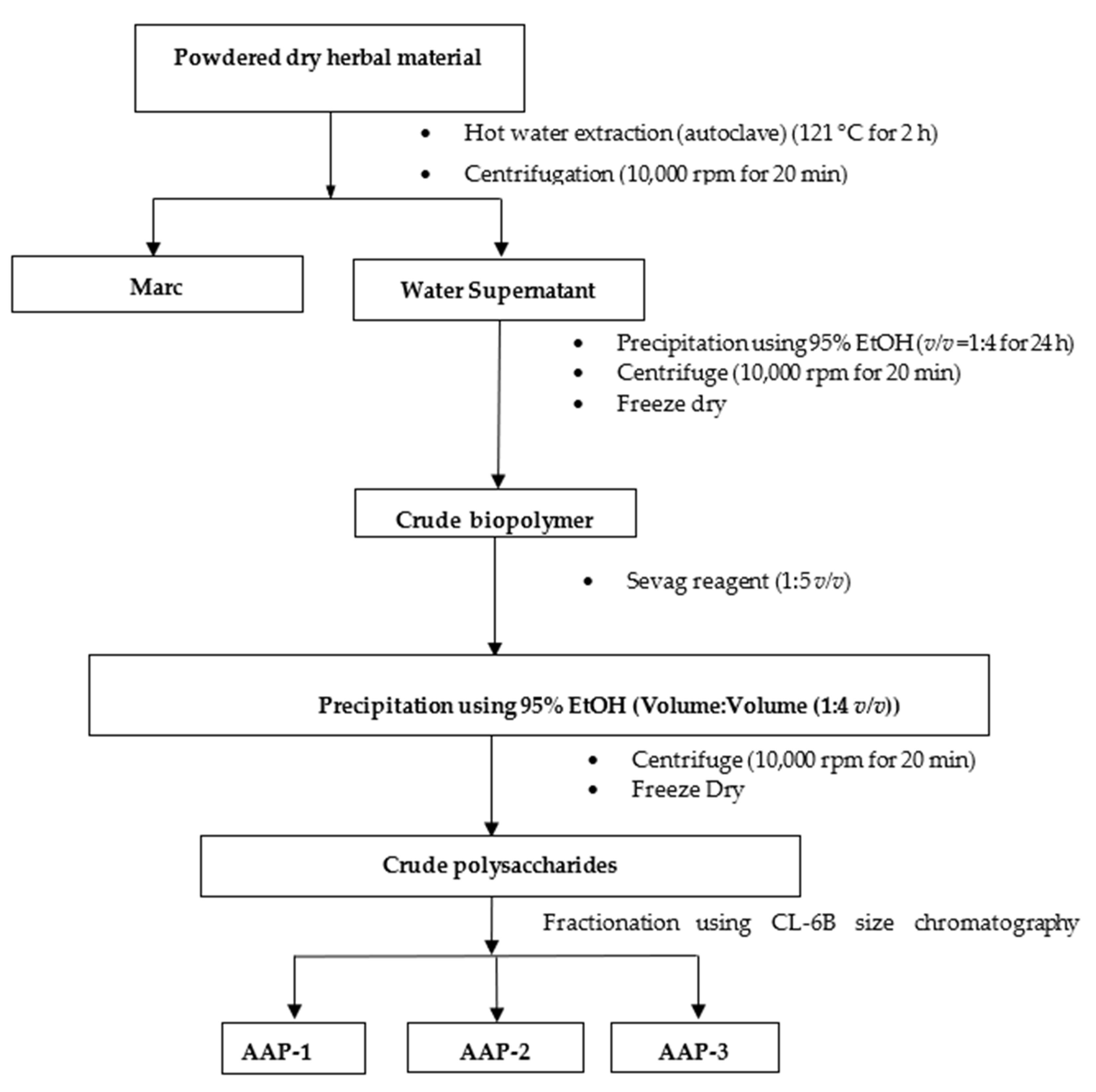
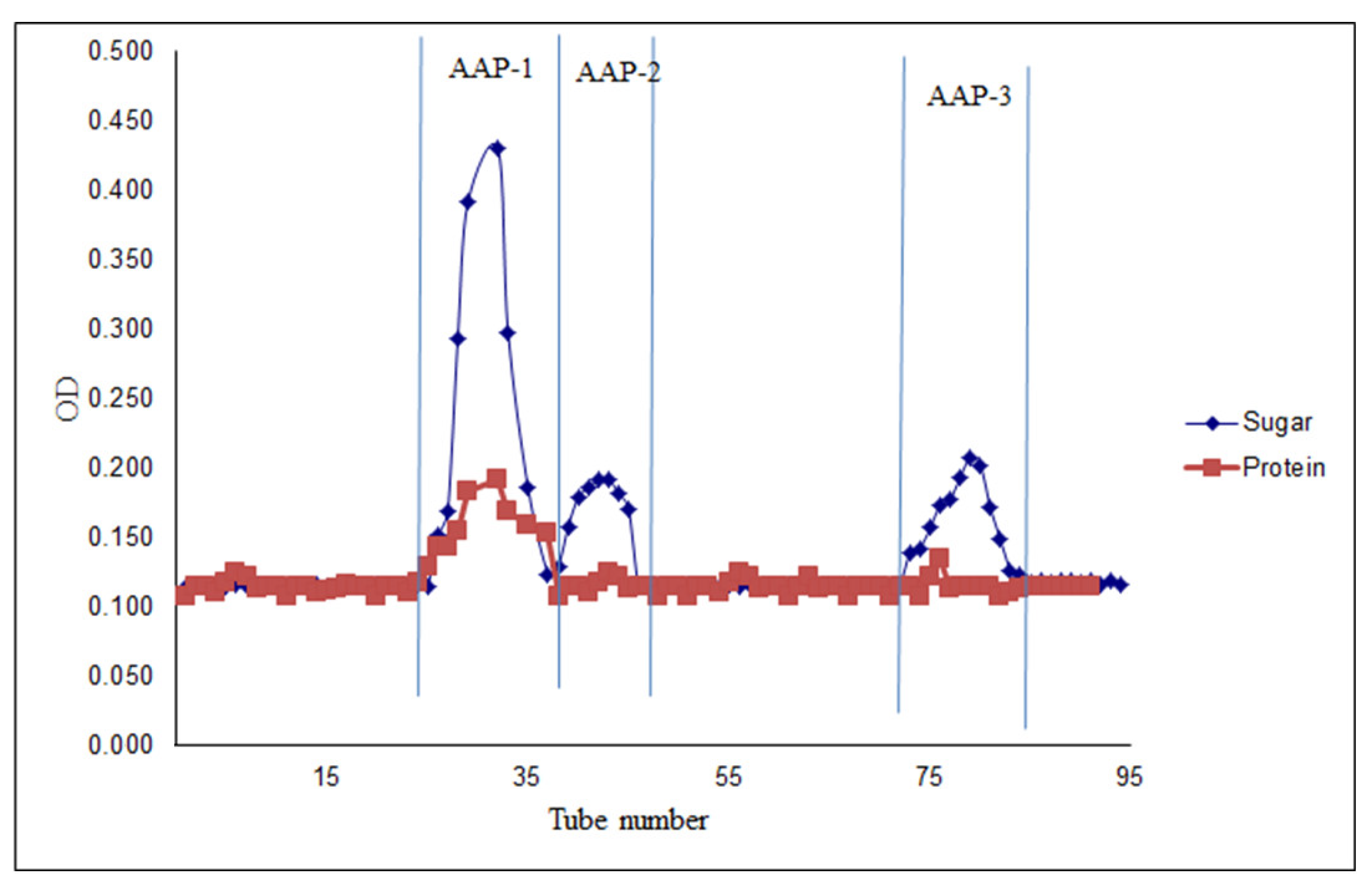
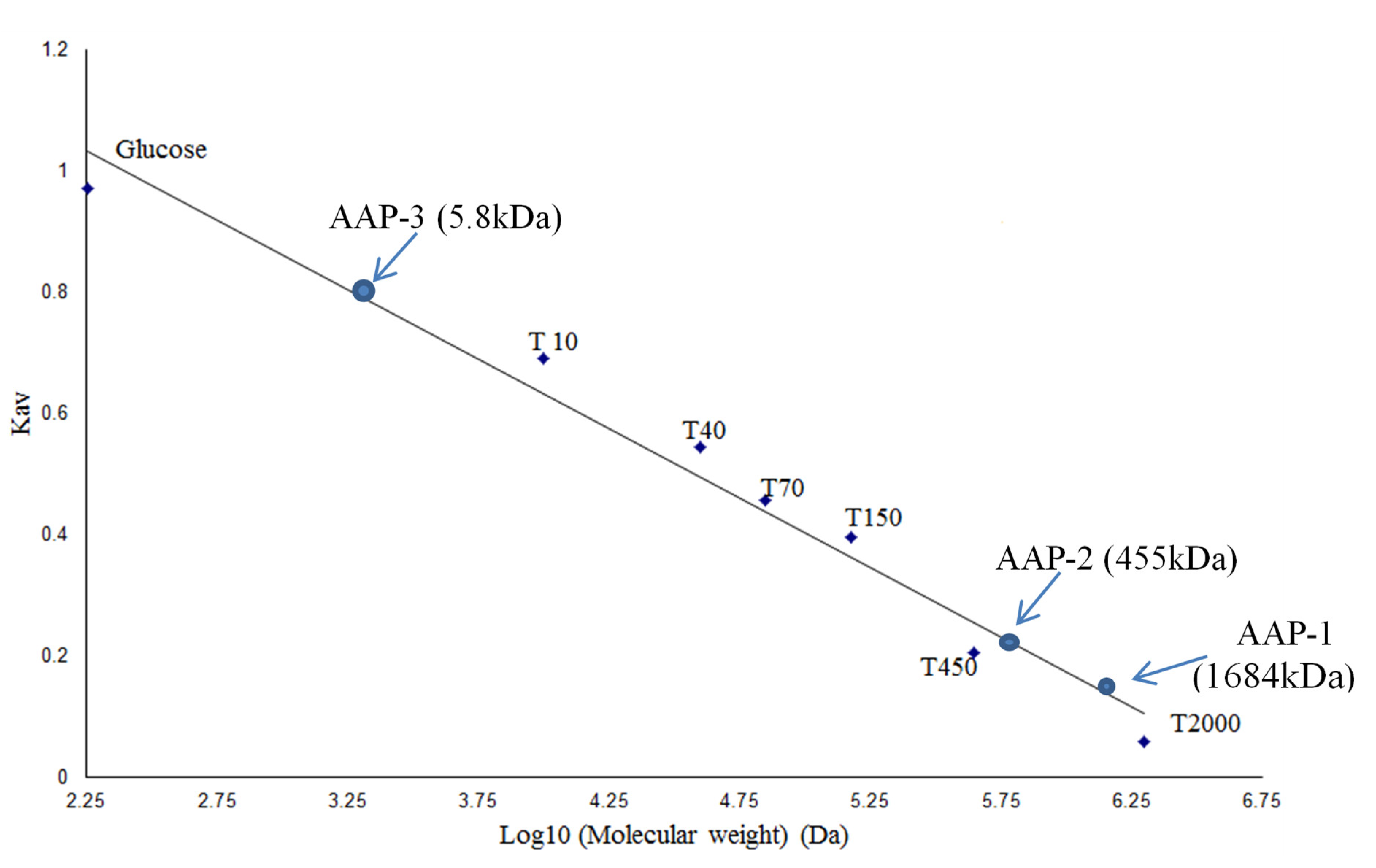
2.1. Fractionation and Purification of Polysaccharides from A. annua
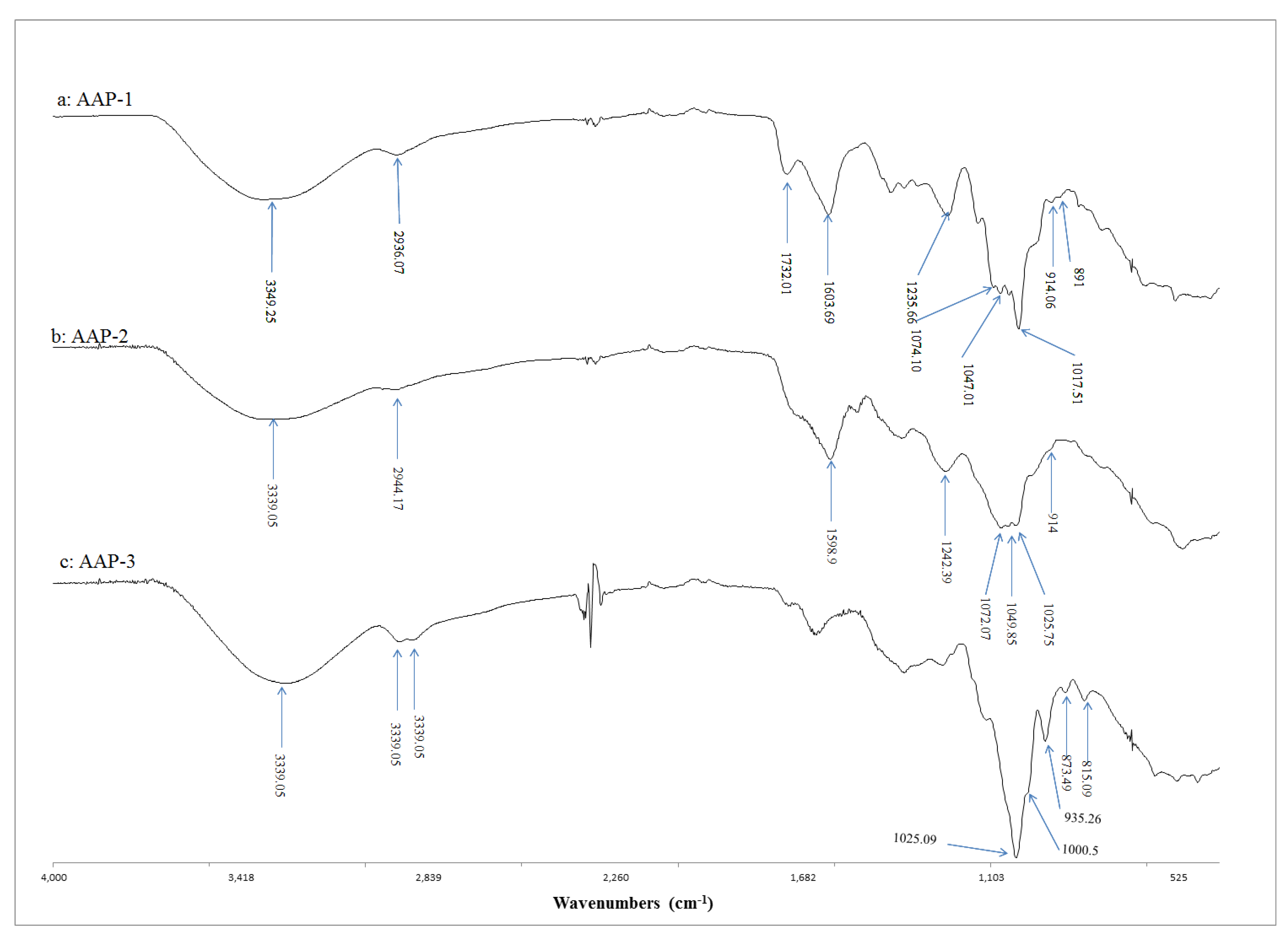
2.2. FTIR Spectroscopic Characterisation of Active Polysaccharides
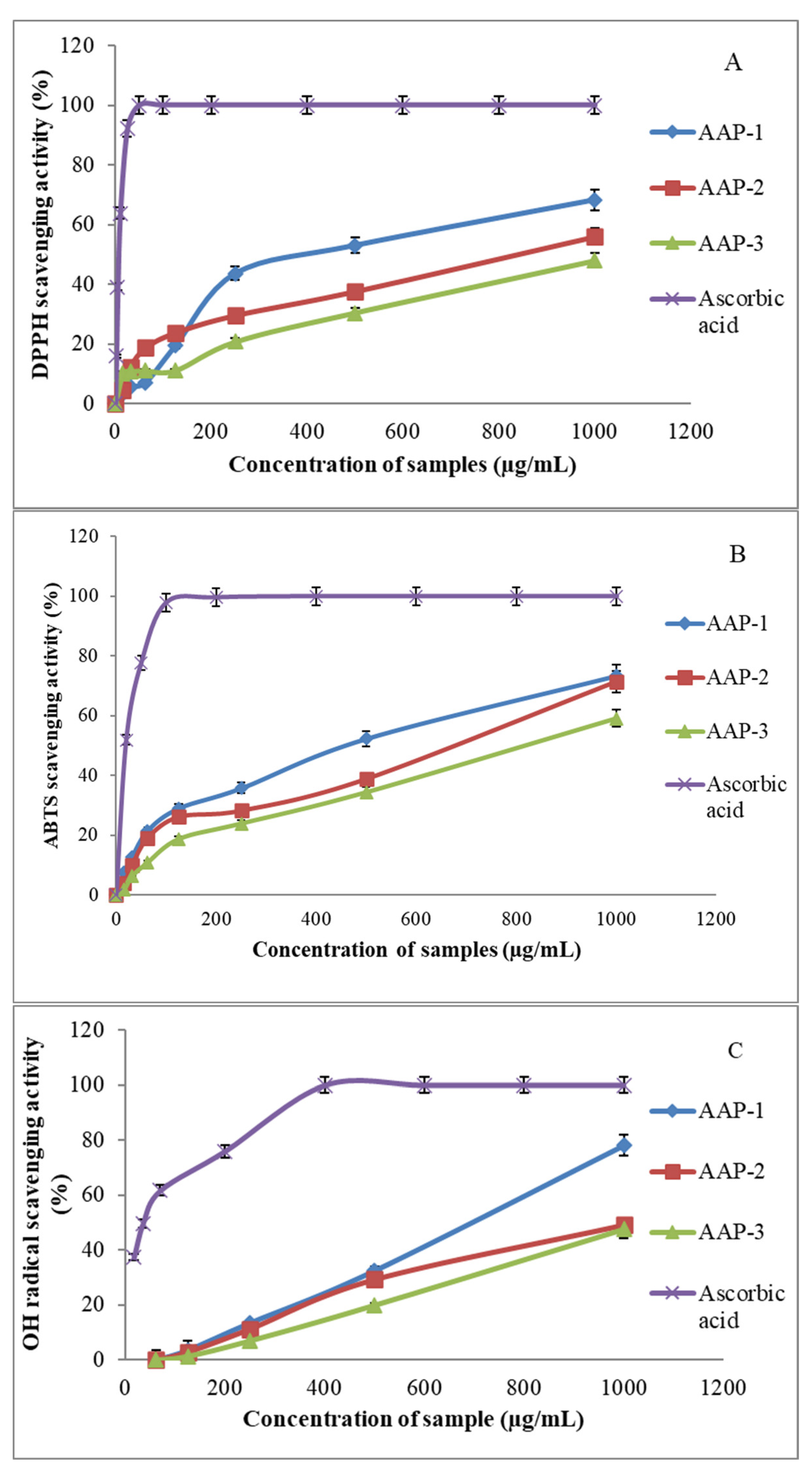
2.3. Antioxidant Activities of AAPs
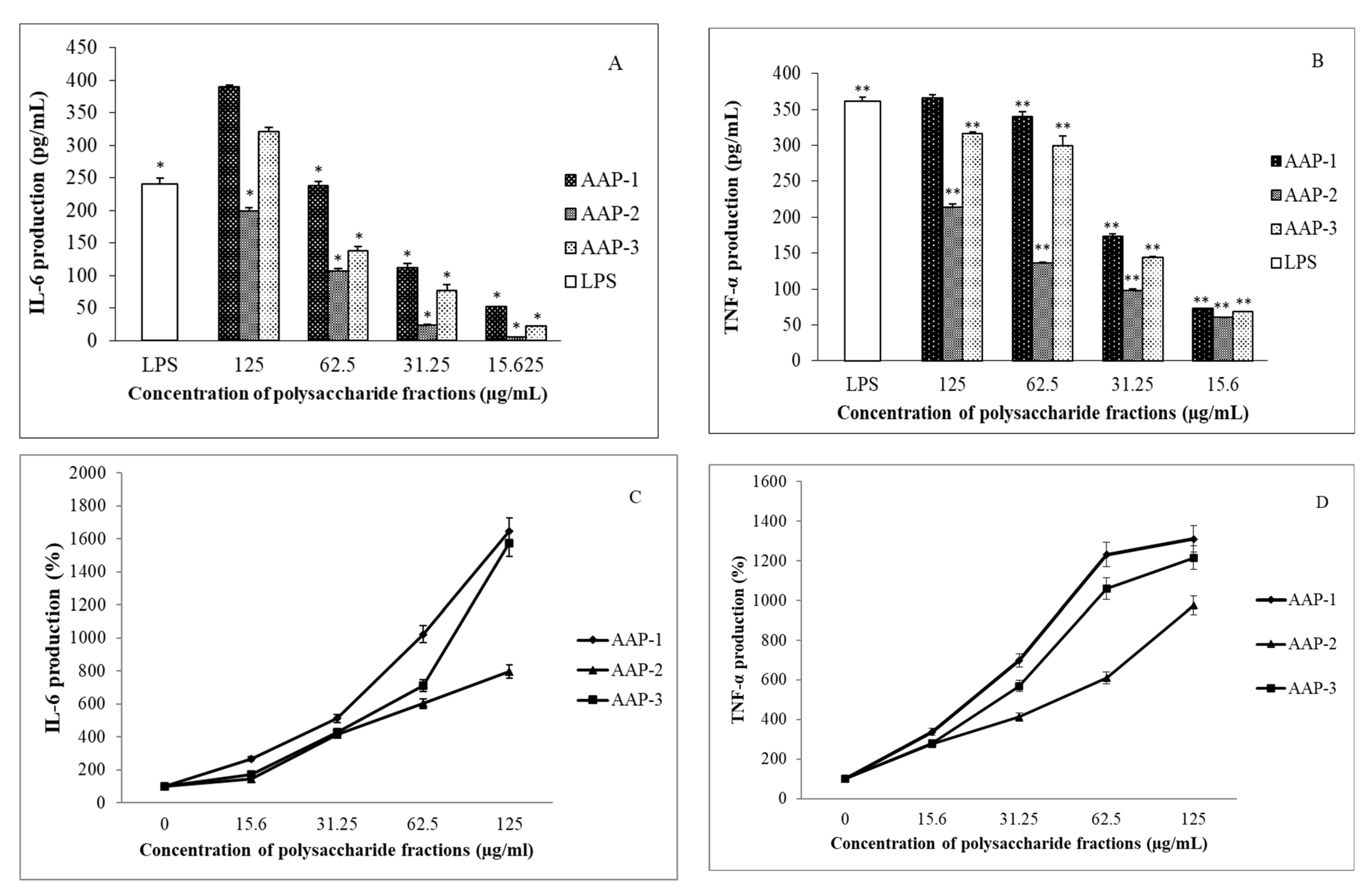
2.4. Immunomodulatory Effects of Polysaccharides from A. annua
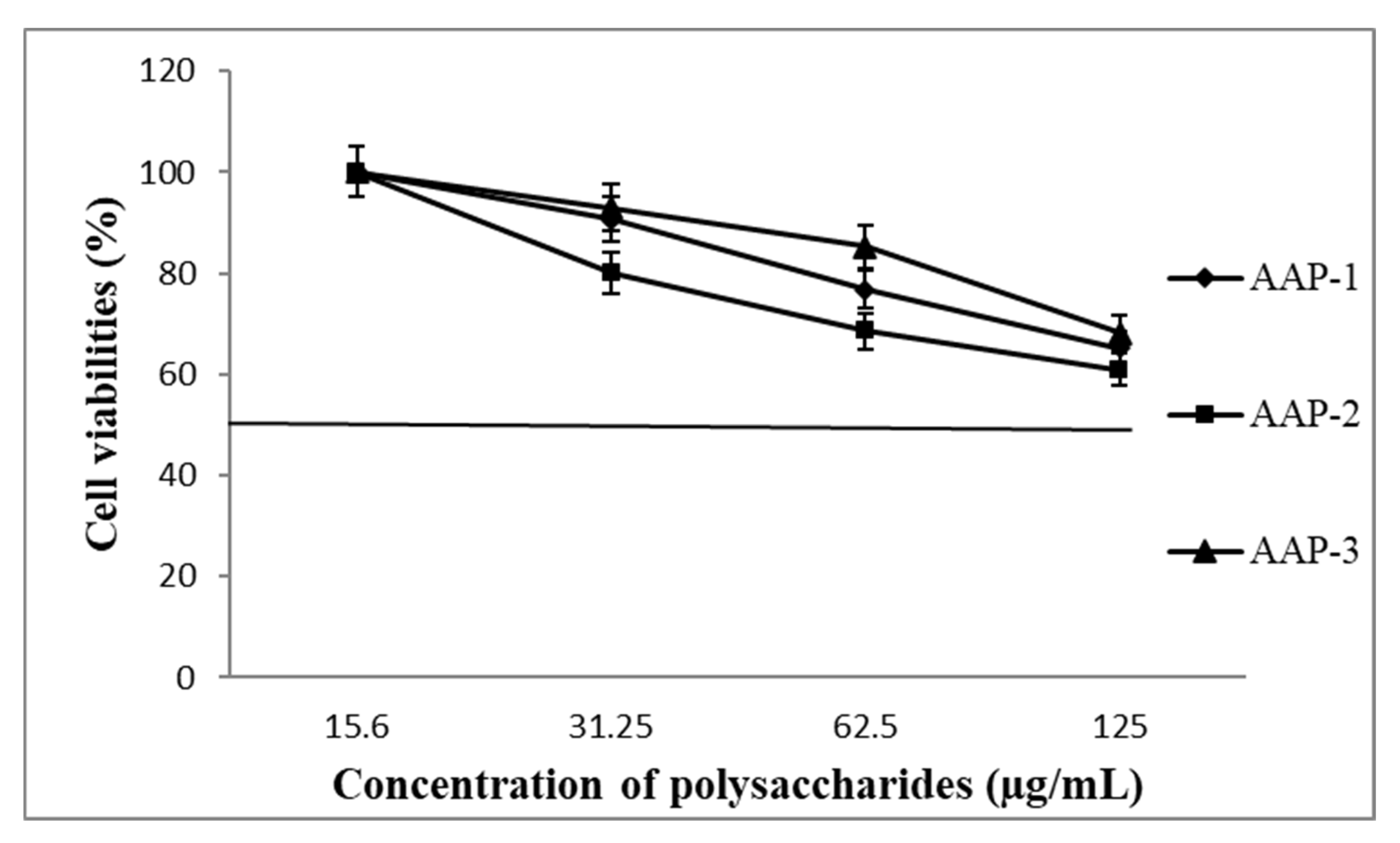
2.5. Cell Viability
3. Materials and Methods
3.1. Material
3.2. Chemicals
3.3. Extraction and Fractionation of Polysaccharides from A. annua
3.4. Determination of Molecular Weights of Polysaccharide Fractions
3.5. Analysis of Monosaccharides
3.6. Bioactivity Tests
3.6.1. DPPH● Scavenging Assay
3.6.2. ABTS●+ Radical Scavenging Assay
3.6.3. OH● Radical Scavenging Assay
3.6.4. Immunomodulatory Activity Assays
IL-6 Production
TNF-α Production
3.6.5. Determination of Cell Viability by MTT Assay
3.7. Fourier Transform Infrared (FTIR) Spectroscopy
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia, L. genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bora, K.S.; Sharma, A. The genus Artemisia: A comprehensive review. Pharm. Biol. 2011, 49, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisibe, E.A.; Umoren, U.E.; Brisibe, F.; Magalhäes, P.M.; Ferreira, J.F.; Luthria, D.; Wu, X.; Prior, R.L. Nutritional characterisation and antioxidant capacity of different tissues of Artemisia annua L. Food Chem. 2009, 115, 1240–1246. [Google Scholar] [CrossRef]
- Tu, Y. From Artemisia Annua L. To Artemisinins. The Discovery and Development of Artemisinins and Antimalarial Agents; Tu, Y., Ed.; Academic Press: London, UK, 2017. [Google Scholar]
- Efferth, T. From ancient herb to versatile, modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017, 46, 65–83. [Google Scholar] [CrossRef]
- Zhang, S.; Mo, Z.; Zhang, S.; Li, X. A network pharmacology approach to reveal the underlying mechanisms of artemisia annua on the treatment of hepatocellular carcinoma. Evid. Based Complementary Altern. Med. 2021, 2021, 8947304. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooy, F.; Sullivan, S.E. The complexity of medicinal plants: The traditional Artemisia annua formulation, current status and future perspectives. J. Ethnopharmacol. 2013, 150, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Hu, Y.; Wang, L.; Ao, C. Ethanol Extract of Artemisia Annua Prevents LPS-Induced Inflammation and Blood–Milk Barrier Disruption in Bovine Mammary Epithelial Cells. Animals 2022, 12, 1228. [Google Scholar] [CrossRef] [PubMed]
- Sankhuan, D.; Niramolyanun, G.; Kangwanrangsan, N.; Nakano, M.; Supaibulwatana, K. Variation in terpenoids in leaves of Artemisia annua grown under different LED spectra resulting in diverse antimalarial activities against Plasmodium falciparum. BMC Plant. Biol. 2022, 22, 1–13. [Google Scholar] [CrossRef]
- Han, X.; Chen, Z.; Yuan, J.; Wang, G.; Han, X.; Wu, H.; Shi, H.; Chou, G.; Yang, L.; Wu, X.J. Artemisia annua water extract attenuates DNCB-induced atopic dermatitis by restraining Th2 cell mediated inflammatory responses in BALB/c mice. J. Ethnopharmacol. 2022, 291, 115160. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Li, J.; Qiu, S.; Qi, F.; Su, H.; Bu, Q.; Jiang, R.; Tang, K.; Zhang, L.; Chen, W.S. The transcription factors TLR1 and TLR2 negatively regulate trichome density and artemisinin levels in Artemisia annua. J. Integr. Plant Biol. 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Zhang, K.; Wang, M.; Qiu, F. Casticin and chrysosplenol D from Artemisia annua L. induce apoptosis by inhibiting topoisomerase IIα in human non-small-cell lung cancer cells. Phytomedicine 2022, 100, 154095. [Google Scholar] [CrossRef]
- Abate, G.; Zhang, L.; Pucci, M.; Morbini, G.; Mac Sweeney, E.; Maccarinelli, G.; Ribaudo, G.; Gianoncelli, A.; Uberti, D.; Memo, M.J.B. Phytochemical Analysis and Anti-Inflammatory Activity of Different Ethanolic Phyto-Extracts of Artemisia annua L. Biomolecules 2021, 11, 975. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, S.S.; Lu, L.; Li, D.; Liang, J.; Huang, Z.H.; Zhou, Y.M.; Zhang, W.J.; Du, S.J. Essential oil from Artemisia annua aerial parts: Composition and repellent activity against two storage pests. Nat. Prod. Res. 2021, 35, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.S.; Huang, Y.; Fidock, D.A.; Polyak, S.J.; Wagoner, J.; Towler, M.J.; Weathers, P.J. Artemisia annua L. extracts inhibit the in vitro replication of SARS-CoV-2 and two of its variants. J. Ethnopharmacol. 2021, 274, 114016. [Google Scholar] [CrossRef]
- Adhikari, B.; Marasini, B.P.; Rayamajhee, B.; Bhattarai, B.R.; Lamichhane, G.; Khadayat, K.; Adhikari, A.; Khanal, S.; Parajuli, N. Potential roles of medicinal plants for the treatment of viral diseases focusing on COVID-19: A review. Phytother. Res. 2021, 35, 1298–1312. [Google Scholar] [CrossRef]
- Elfawal, M.A.; Gray, O.; Dickson-Burke, C.; Weathers, P.J.; Rich, S.M. Artemisia annua and artemisinins are ineffective against human Babesia microti and six Candida sp. Longhua Chin. Med. 2021, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Shankar, G.; Mukhopadhyay, P.; Gupta, V. A concise review on Artemisia annua L.: A major source of diverse medicinal compounds. Ind. Crops. Prod. 2022, 184, 115072. [Google Scholar] [CrossRef]
- Cheng, K.F.; Leung, P.C. General review of polysaccharopeptides (PSP) from C. versicolor: Pharmacological and clinical studies. Cancer Ther. 2008, 6, 117–130. [Google Scholar]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Zhang, L.; Koyyalamudi, S.R.; Reddy, N. Isolation, Characterization, and Biological Activities of Polysaccharides from Medicinal Plants and Mushrooms. In Studies in Nature Products Chemistry, 1st ed.; Atta-Ur-Rahman, F.R.S., Ed.; Elsevier: London, UK, 2014; pp. 117–147. [Google Scholar]
- Zhang, L.; Reddy, N. Bioactive Molecules from Medicinal Herbs for Life Threating Diseases. Int. J. Mol. Sci. 2018, 2, 4. [Google Scholar]
- Friedman, M. Mushroom Polysaccharides: Chemistry and Antiobesity, Antidiabetes, Anticancer, and Antibiotic Properties in Cells, Rodents, and Humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, Y. Polysaccharides. In Immunotherapy of Cancer; Yamaguchi, Y., Ed.; Springer: Tokyo, Japan, 2016; pp. 37–50. [Google Scholar]
- Zhang, L.; Li, C.G.; Liang, H.; Reddy, N. Bioactive Mushroom Polysaccharides: Immunoceuticals to Anticancer Agents. J. Nutraceuticals Food Sci. 2017, 2, 6. [Google Scholar]
- Huo, J.; Lu, Y.; Xia, L.; Chen, D. Structural characterization and anticomplement activities of three acidic homogeneous polysaccharides from Artemisia annua. J. Ethnopharmacol. 2020, 247, 112281. [Google Scholar] [CrossRef]
- Yan, L.; Xiong, C.; Xu, P.; Zhu, J.; Yang, Z.; Ren, H.; Luo, Q. Structural characterization and in vitro antitumor activity of A polysaccharide from Artemisia annua L. (Huang Huahao). Carbohydr. Polym. 2019, 213, 361–369. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Liu, C. Anti-tumour effects of polysaccharides isolated from Artemisia Annua L by inducing cell apoptosis and immunomodulatory anti-hepatoma effects of polysaccharides. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Lin, D.; Alim, A.; Zheng, Q.; Yang, X. Chemical characterization of a novel polysaccharide ASKP-1 from Artemisia sphaerocephala Krasch seed and its macrophage activation via MAPK, PI3k/Akt and NF-κBsignaling pathways in RAW264. 7 cells. Food Funct. 2017, 8, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Shi, C.; Shi, S.; Wang, H.; Yan, J.; Wang, S. An inulin-type fructan isolated from Artemisia japonica and its anti-arthritic effects. J. Funct. Foods 2017, 29, 29–36. [Google Scholar] [CrossRef]
- Zhang, L.; Khoo, C.S.; Koyyalamudi, S.R.; Jones, E.C.; Reddy, N. Immunostimulatory and anticancer activities of polysaccharides extracted from traditional anticancer Chinese medicinal herbs. Pharmacologia 2018, 9, 18–29. [Google Scholar]
- Zhang, L.; Khoo, C.S.; Koyyalamudi, S.R.; Jones, E.C.; Reddy, N. Antioxidant and immunomodulatory activities and structural characterization of polysaccharides isolated from Lobelia chinensis Lour. Pharmacologia 2018, 9, 157–168. [Google Scholar]
- Fan, H.; Liu, F.; Bligh, S.A.; Shi, S.; Wang, S. Structure of a homofructosan from Saussureacostus and anti-complementary activity of its sulfated derivatives. Carbohydr. Polym. 2014, 105, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Antioxidant activities and characterisation of polysaccharides isolated from the seeds of Lupinus angustifolius. Ind. Crops Prod. 2015, 74, 950–956. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, L.M. Chemical structural and chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Polym. 2009, 76, 349–361. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.; Liu, M.; Wang, Q.; Li, Y. Extraction optimization, characterization, antioxidant and immunomodulatory activities of a novel polysaccharide from the wild mushroom Paxillus involutus. Int. J. Biol. Macromol. 2018, 112, 326–332. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.M.; Qi, Z.M.; Wang, S.Y.; Liu, S.X.; Li, X.; Wang, H.J.; Xia, X.C. An overview on natural polysaccharides with antioxidant properties. Curr. Med. Chem. 2013, 20, 2899–2913. [Google Scholar] [CrossRef]
- Jiang, M.H.; Zhu, L.; Jiang, J.G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef] [PubMed]
- Peshev, D.; Van den Ende, W. Fructans: Prebiotics and immunomodulators. J. Funct. Foods 2014, 8, 348–357. [Google Scholar] [CrossRef]
- Franco-Robles, E.; López, M.G. Implication of fructans in health: Immunomodulatory and antioxidant mechanisms. Sci. World J. 2015, 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Vogt, L.; Ramasamy, U.; Meyer, D.; Pullens, G.; Venema, K.; Faas, M.M.; Schols, H.A.; de Vos, P. Immune modulation by different types of β2→1-fructans is toll-like receptor dependent. PLoS ONE 2013, 8, e68367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [Green Version]
- de Avellar, I.G.; Magalhães, M.M.; Silva, A.B.; Souza, L.L.; Leitão, A.C.; Hermes-Lima, M. Re-evaluating the role of 1, 10-phenanthroline in oxidative reactions involving ferrous ions and DNA damage. Biochim. Biophys. Acta BBA Gen. Subjects 2004, 1675, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.J.; Wang, N.N.; Zhang, L.G.; Guo, Y.Z.; Shi, W.Z. Evaluation of the effects of active fractions of chinese medicine formulas on IL-1β, IL-6, and TNF-α release from ANA-1 murine macrophages. J. Ethnopharmacol. 2016, 179, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Wu, J.; Zhao, M.; Sun, W.; Sun, J.; Li, H.; Huang, M. Immunomodulatory activity of a novel polysaccharide extracted from Huangshui on THP-1 cells through NO production and increased IL-6 and TNF-α expression. Food Chem. 2020, 330, 127257. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AAP-1 | AAP-2 | AAP-3 * | |
|---|---|---|---|
| Protein (%) | 29.57 | 10.95 | 9.32 |
| Carbohydrate (%) | 70.43 | 89.05 | 90.68 |
| Monosaccharide (% ratio) | |||
| Rhamnose (%) | 9.67 | ||
| Ribose (%) | |||
| Fucose (%) | |||
| Arabinose (%) | 33.35 | 36.02 | |
| Xylose (%) | 7.67 | ||
| Mannose (%) | 1.44 | 18.17 | 53.09 |
| Galactose (%) | 30.92 | 16.07 | |
| Glucose (%) | 8.69 | 22.16 | 46.92 |
| Unknown (%) | 8.26 | 7.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Reddy, N.; Khoo, C.S.; Koyyalamudi, S.R. Structural Characterization and In-Vitro Antioxidant and Immunomodulatory Activities of Polysaccharide Fractions Isolated from Artemisia annua L. Molecules 2022, 27, 3643. https://doi.org/10.3390/molecules27113643
Zhang L, Reddy N, Khoo CS, Koyyalamudi SR. Structural Characterization and In-Vitro Antioxidant and Immunomodulatory Activities of Polysaccharide Fractions Isolated from Artemisia annua L. Molecules. 2022; 27(11):3643. https://doi.org/10.3390/molecules27113643
Chicago/Turabian StyleZhang, Lin, Narsimha Reddy, Cheang Soo Khoo, and Sundar Rao Koyyalamudi. 2022. "Structural Characterization and In-Vitro Antioxidant and Immunomodulatory Activities of Polysaccharide Fractions Isolated from Artemisia annua L." Molecules 27, no. 11: 3643. https://doi.org/10.3390/molecules27113643