
Fecal Metabolic Profiling of Breast Cancer Patients during Neoadjuvant Chemotherapy Reveals Potential Biomarkers

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
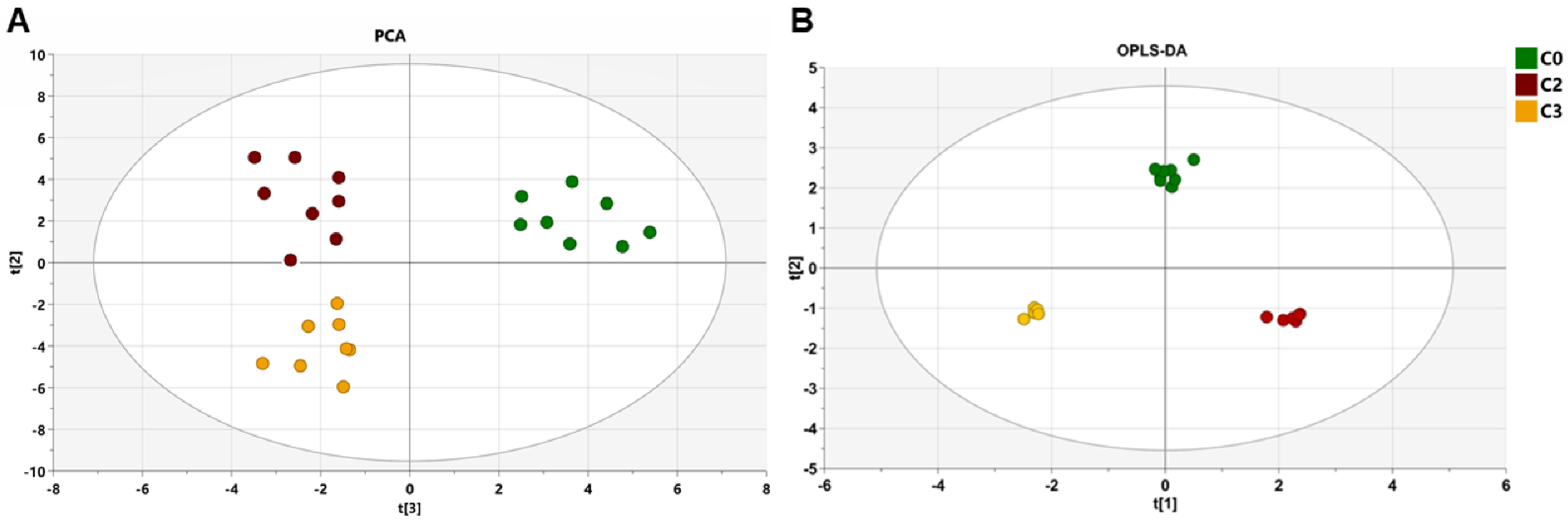
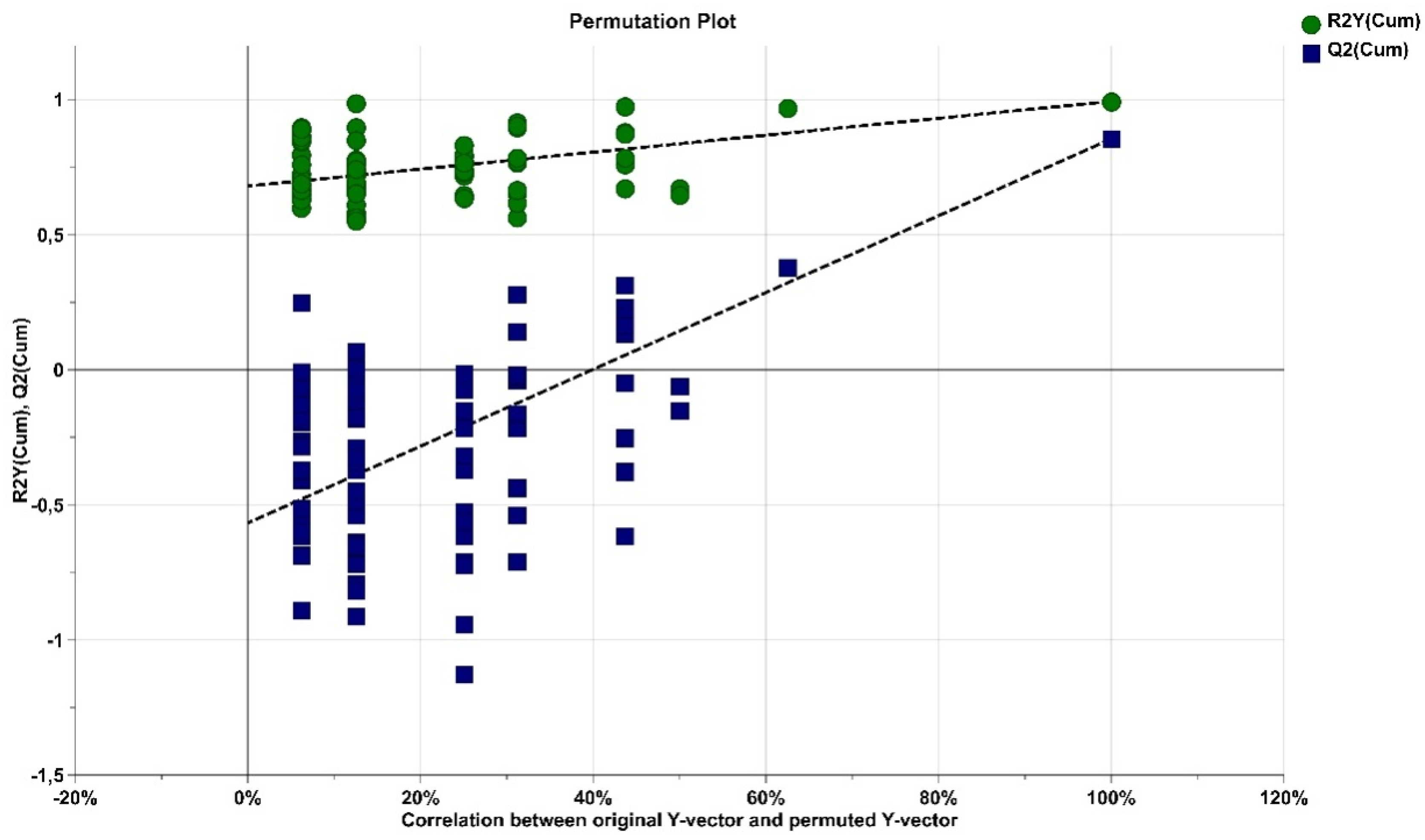
2.2. Impact of Three Chemotherapy Cycles on the Differential Fecal Metabolites of Breast Cancer Patients
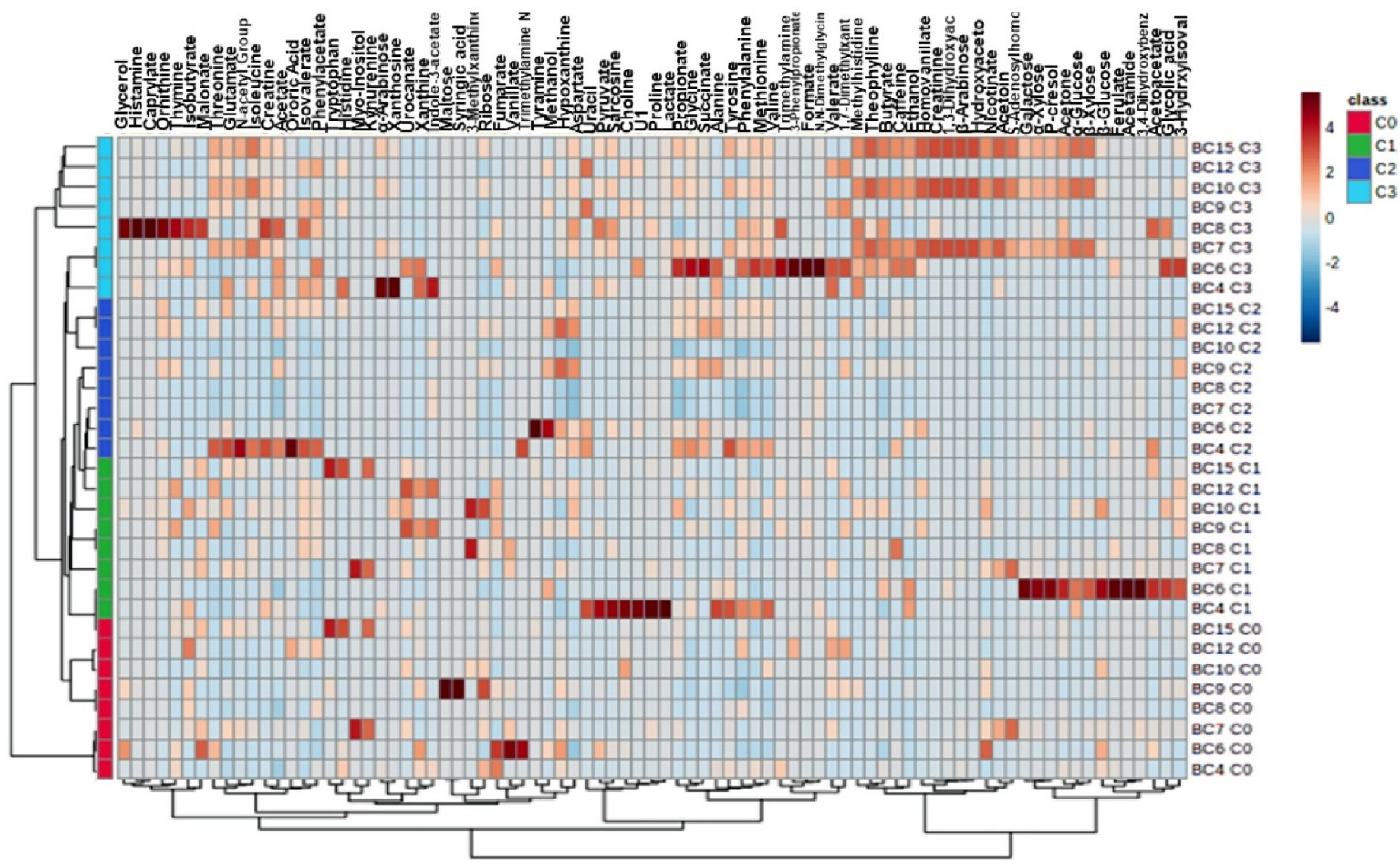
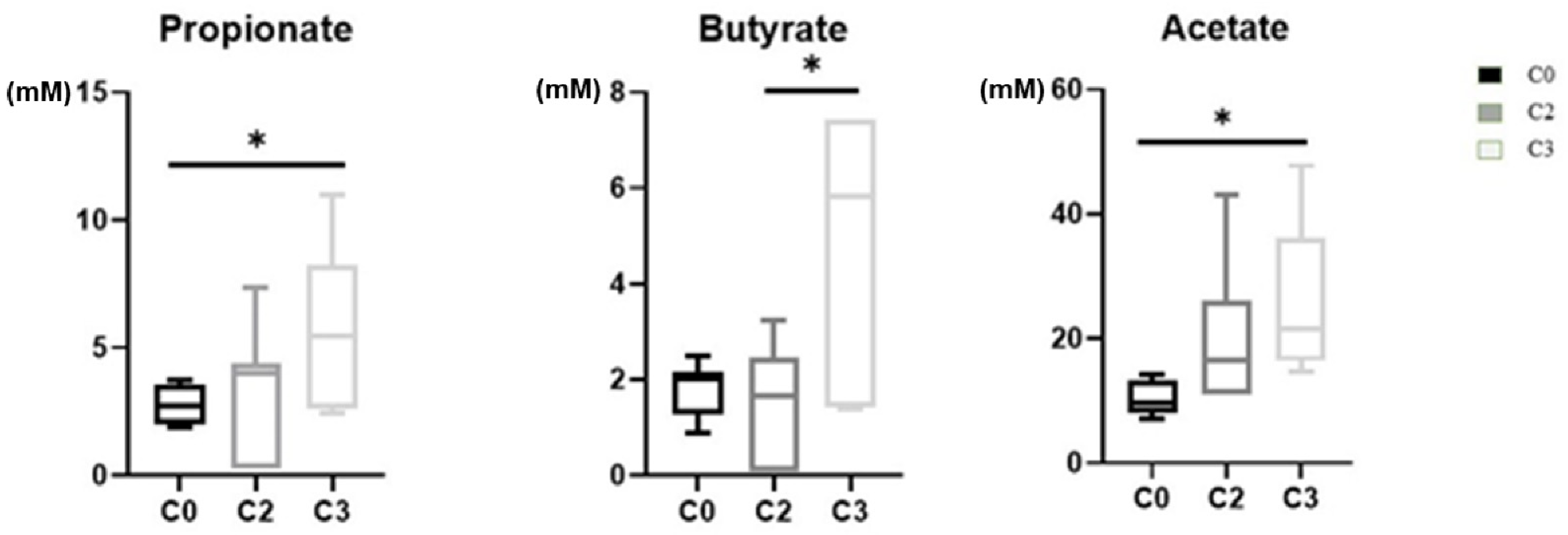
2.3. Changes of Metabolite Levels in Breast Cancer Patients before and after Chemotherapy
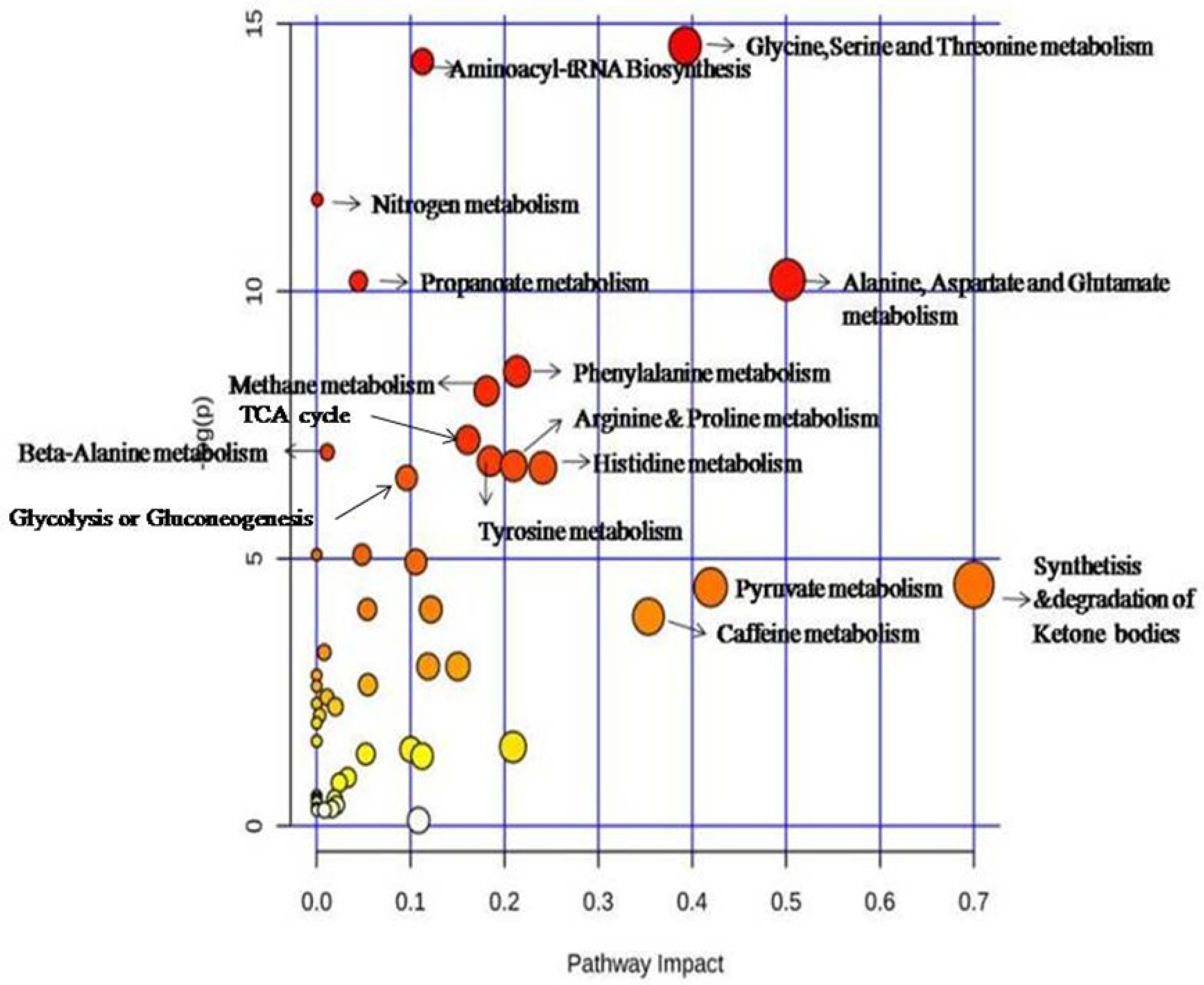
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Fecal Metabolite Extraction
4.3. NMR Spectroscopy
4.4. NMR Data Processing
4.5. Metabolite Identification
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- World Health Organization. “Latest Global Cancer Data: Cancer Burden Rises to 18.1 million New Cases and 9.6 million Cancer Deaths in 2018.” International Agency for Research on Cancer. Geneva: World Health Organization (2018). Available online: http://gco.iarc.fr/ (accessed on 12 September 2018).
- Malvezzi, M.; Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E. European cancer mortality predictions for the year 2019 with focus on breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2019, 30, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Mejri, N.; Boussen, H.; Labidi, S.; Bouzaiene, H.; Afrit, M.; Benna, F.; Rahal, K. Inflammatory breast cancer in Tunisia from 2005 to 2010: Epidemiologic and anatomoclinical transitions from published data. Asian Pac. J. Cancer Prev. 2015, 16, 1277–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austreid, E.; Lonning, P.E.; Eikesdal, H.P. The emergence of targeted drugs in breast cancer to prevent resistance to endocrine treatment and chemotherapy. Expert Opin. Pharmacother. 2014, 15, 681–700. [Google Scholar] [CrossRef]
- Kim, I.S.; Gao, Y.; Welte, T.; Wang, H.; Liu, J.; Janghorban, M.; Sheng, K.; Niu, Y.; Goldstein, A.; Zhao, N.; et al. Immuno-subtyping of breast cancer reveals distinct myeloid cell profiles and immunotherapy resistance mechanisms. Nat. Cell Biol. 2019, 21, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Pajic, M.; Froio, D.; Daly, S.; Doculara, L.; Millar, E.; Graham, P.H.; Drury, A.; Steinmann, A.; de Bock, C.E.; Boulghourjian, A.; et al. miR-139-5p modulates radiotherapy resistance in breast cancer by repressing multiple gene networks of DNA repair and ROS defense. Cancer Res. 2018, 78, 501–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foukakis, T.; von Minckwitz, G.; Bengtsson, N.O.; Brandberg, Y.; Wallberg, B.; Fornander, T.; Mlineritsch, B.; Schmatloch, S.; Singer, C.F.; Steger, G.; et al. Effect of tailored dose-dense chemotherapy vs standard 3-weekly adjuvant chemotherapy on recurrence-free survival amongwomen with high-risk early breast cancer: A randomized clinical trial. JAMA J. Am. Med. Assoc. 2016, 316, 1888–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulombe, G.; Tyldesley, S.; Speers, C.; Paltiel, C.; Aquino-Parsons, C.; Bernstein, V.; Truong, P.T.; Keyes, M.; Olivotto, I.A. Is Mastectomy Superior to Breast-Conserving Treatment for Young Women? Int. J. Radiat. Oncol. Biol. Phys. 2007, 67, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef] [PubMed]
- Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Commentary. Metabolic profiling: A Rosetta Stone for genomics? Curr. Opin. Plant Biol. 1999, 2, 83–85. [Google Scholar] [CrossRef]
- Corona, G.; Rizzolio, F.; Giordano, A.; Toffoli, G. Pharmaco-metabolomics: An emerging “omics” tool for the personalization of anticancer treatments and identification of new valuable therapeutic targets. J. Cell Physiol. 2012, 227, 2827–2831. [Google Scholar] [CrossRef]
- Wei, S.; Liu, L.; Zhang, J.; Bowers, J.; Gowda, G.N.; Seeger, H.; Fehm, T.; J.Neubauer, H.; Vogel, U.; E. Clare, S.; et al. Metabolomics approach for predicting response to neoadjuvant chemotherapy for breast cancer. Mol. Oncol. 2013, 7, 297–307. [Google Scholar] [CrossRef] [Green Version]
- McCartney, A.; Vignoli, A.; Biganzoli, L.; Love, R.; Tenori, L.; Luchinat, C.; Di Leo, A. Metabolomics in breast cancer: A decade in review. Cancer Treat. Rev. 2018, 67, 88–96. [Google Scholar] [CrossRef]
- Miolo, G.; Muraro, E.; Caruso, D.; Crivellari, D.; Ash, A.; Scalone, S.; Lombardi, D.; Rizzolio, F.; Giordano, A.; Corona, G. Phamacometabolomics study identifies circulating spermidine and tryptophan as potential biomarkers associated with the complete pathological response to trastuzumab-paclitaxel neoadjuvant therapy in HER-2 positive breast cancer. Oncotarget 2016, 7, 39809–39822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.S.; Baek, H.M.; Kim, S.; Jung Kim, M.; Youk, J.H.; Moon, H.J.; Kim, E.K.; Nam, Y.K. Magnetic resonance metabolic profiling of breast cancer tissue obtained with core needle biopsy for predicting pathologic response to neoadjuvant chemotherapy. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Baek, H.M.; Chen, J.H.; Nie, K.; Yu, H.J.; Bahri, S.; Mehta, R.S.; Nalcioglu, O.; Su, M.Y. Predicting pathologic response to neoadjuvant chemotherapy in breast cancer by using MR imaging and quantitative 1H MR spectroscopy. Radiology 2009, 251, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Jiang, C.; Huang, S.; Gong, X.; Wang, S.; Shen, P. Analysis of urinary metabolites for breast cancer patients receiving chemotherapy by CE-MS coupled with on-line concentration. Clin. Biochem. 2013, 46, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yan, L.; Liu, S.; Ambrosone, C.B.; Zhao, H. Plasma metabolomic profiles in breast cancer patients and healthy controls: By race and tumor receptor subtypes. Transl. Oncol. 2013, 6, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.; Yang, T. Identification of possible biomarkers for breast cancer from free fatty acid profiles determined by GC-MS and multivariate statistical analysis. Clin. Biochem. 2012, 45, 127–133. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Zhang, X. Metabonomics studies on serum and urine of patients with breast cancer using 1H-NMR spectroscopy. Oncotarget 2017, 1–9. Available online: https://www.oncotarget.com/article/16210/ (accessed on 13 April 2021).
- Chae, E.Y.; Shin, H.J.; Kim, S.; Baek, H.M.; Yoon, D.; Kim, S.; Shim, Y.E.; Kim, H.H.; Cha, J.H.; Choi, W.J.; et al. The role of high-resolution magic angle spinning 1H nuclear magnetic resonance spectroscopy for predicting the invasive component in patients with ductal carcinoma in situ diagnosed on preoperative biopsy. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef]
- Rakha, E.A.; Chan, S. Metastatic Triple-negative Breast Cancer. Clin. Oncol. 2011, 23, 587–600. [Google Scholar] [CrossRef]
- Weir, T.L.; Manter, D.K.; Sheflin, A.M.; Barnett, B.A.; Heuberger, A.L.; Ryan, E.P. Stool Microbiome and Metabolome Differences between Colorectal Cancer Patients and Healthy Adults. PLoS ONE 2013, 8, e70803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, F.; Adewiah, S.; Syam, A.F.; Fatchiyah, F. Altered profile of gut microbiota and the level short chain fatty acids in colorectal cancer patients. J. Phys. Conf. Ser. 2019, 1146, 012037. [Google Scholar] [CrossRef]
- Weickert, M.O.; Pfeiffer, A.F.H. Metabolic Effects of Dietary Fiber Consumption and Prevention of Diabetes. J. Nutr. 2008, 138, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8836–8847. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, G. Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef]
- Panebianco, C.; Andriulli, A.; Pazienza, V. Pharmacomicrobiomics: Exploiting the drug-microbiota interactions in anticancer therapies. Microbiome 2018, 6, 92. [Google Scholar] [CrossRef]
- Lehouritis, P.; Cummins, J.; Stanton, M.; Murphy, C.T.; McCarthy, F.O.; Reid, G.; Urbaniak, C.; Byrne, W.L.; Tangney, M. Local bacteria affect the efficacy of chemotherapeutic drugs. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Goubet, A.G.; Daillère, R.; Routy, B.; Derosa, L.; MRoberti, P.; Zitvogel, L. The impact of the intestinal microbiota in therapeutic responses against cancer. Comptes Rendus Biol. 2018, 341, 284–289. [Google Scholar] [CrossRef]
- Ma, W.; Mao, Q.; Xia, W.; Dong, G.; Yu, C.; Jiang, F. Gut microbiota shapes the efficiency of cancer therapy. Front. Microbiol. 2019, 10, 1050. [Google Scholar] [CrossRef] [Green Version]
- Zitvogel, L.; Galluzzi, L.; Viaud, S.; Vétizou, M.; Daillère, R.; Merad, M.; Kroemer, G. Cancer and the gut microbiota: An unexpected link. Sci. Transl. Med. 2015, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, F.; Linsalata, M.; Clemente, C.; D’Attoma, B.; Orlando, A.; Campanella, G.; Giotta, F.; Riezzo, G. The effects of fluorouracil, epirubicin, and cyclophosphamide (FEC60) on the intestinal barrier function and gut peptides in breast cancer patients: An observational study. BMC Cancer 2013, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, L.; Zunino, F. Overview of tumor cell chemoresistance mechanisms. Methods Mol. Med. 2005, 11, 127–148. [Google Scholar]
- Therasse, P.; Arbuck, S.G.; Eisenhauer, E.A.; Wanders, J.; Kaplan, R.S.; Rubinstein, L.; Verweij, J.; Van Glabbeke, M.; Van Oosterom, A.T.; Christian, M.C.; et al. New guidelines to evaluate the response to treatment in solid tumors. J. Natl. Cancer Inst. 2000, 92, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, A.P.; Redinbo, M.R.; Bultman, S.J. The role of the microbiome in cancer development and therapy. CA Cancer J. Clin. 2017, 67, 326–344. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef]
- Eniu, D.T.; Romanciuc, F.; Moraru, C.; Goidescu, I.; Eniu, D.; Staicu, A.; Rachieriu, C.; Buiga, R.; Socaciu, C. The decrease of some serum free amino acids can predict breast cancer diagnosis and progression. Scand. J. Clin. Lab. Investig. 2019, 79, 17–24. [Google Scholar] [CrossRef]
- Mishra, P.; Ambs, S.; Mishra, P.; Ambs, S. Metabolic signatures of human breast cancer Metabolic signatures of human breast cancer. Mol. Cell Oncol. 2015, 2, e992217. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite Profiling Identifies a Key Role for Glycine in Rapid Cancer Cell Proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.C.; Ng, S.C.; Yang, H.; Rai, A.; Umashankar, S.; Ma, S.; Soh, B.S.; Sun, L.L.; Tai, B.C.; Nga, M.E.; et al. Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis. Cell 2012, 148, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Possemato, R.; Marks, K.M.; Shaul, Y.D.; Pacold, M.E.; Kim, D.; Birsoy, K.; Sethumadhavan, S.; Woo, H.K.; Jang, H.G.; Jha, A.K.; et al. Functional genomics reveal that the serine synthesis pathway is essential in breast cancer. Nature 2011, 476, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Zhou, B.; Zhao, A.; Qiu, Y.; Zhao, X. Lowered circulating aspartate is a metabolic feature of human breast cancer. Oncotarget 2015, 6, 33369–33381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, D.A.; Winnike, J.H.; McRitchie, S.L.; Clark, R.F.; Pathmasiri, W.W.; Sumner, S.J. Metabolomics Analysis of Hormone-Responsive and Triple-Negative Breast Cancer Cell Responses to Paclitaxel Identify Key Metabolic Differences. J. Proteome Res. 2016, 15, 3225–3240. [Google Scholar] [CrossRef]
- Walenta, S.; Mueller-Klieser, W.F. Lactate: Mirror and motor of tumor malignancy. Semin. Radiat. Oncol. 2004, 14, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Walenta, S.; Wetterling, M.; Lehrke, M.; Schwickert, G.; Sundfør, K.; Rofstad, E.K.; Mueller-Klieser, W. High lactate levels predict likelihood of metastases, tumor recurrence, and restricted patient survival in human cervical cancers. Cancer Res. 2000, 60, 916–921. [Google Scholar]
- Hodge, C.J.; Apkarian, A.V. The Metabolic Requirements of Cell Proliferation. Science 1990, 5, 363–397. [Google Scholar]
- Razungles, J.; Cavaillès, V.; Jalaguier, S.; Teyssier, C. L’effet Warburg. Med. Sci. 2013, 29, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Sermonti, G. On the origin of the origin. Riv. Biol. 2009, 102, 309–314. [Google Scholar]
- Sidney, W.H.; Warburg, O.; Dean, B.; Arthur, L.S. On Respiratory Impairment in Cancer Cells. Am. Assoc. Adv. Sci. 1956, 124, 267–272. Available online: https://www.jstor.org/stable/1751794 (accessed on 10 August 2020).
- Ryu, C.S.; Kwak, H.C.; Lee, K.S.; Kang, K.W.; Oh, S.J.; Lee, K.H.; Kim, H.M.; Ma, J.Y.; Kim, S.K. Sulfur amino acid metabolism in doxorubicin-resistant breast cancer cells. Toxicol. Appl. Pharmacol. 2011, 255, 94–102. [Google Scholar] [CrossRef]
- Euceda, L.R.; Haukaas, T.H.; Giskeødegård, G.F.; Vettukattil, R.; Engel, J.; Silwal-Pandit, L.; Silwal-Pandit, L.; Lundgren, S.; Borgen, E.; Garred, Ø.; et al. Evaluation of metabolomic changes during neoadjuvant chemotherapy combined with bevacizumab in breast cancer using MR spectroscopy. Metabolomics 2017, 13, 37. [Google Scholar] [CrossRef]
- Fuhler, G.M.; Eppinga, H.; Peppelenbosch, M.P. Fumarates and Cancer. Trends Mol. Med. 2017, 23, 3–5. [Google Scholar] [CrossRef]
- Sciacovelli, M.; Frezza, C. Oncometabolites: Unconventional triggers of oncogenic signalling cascades. Free Radic. Biol. Med. 2016, 100, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspuria, P.J.P.; Lunt, S.Y.; Väremo, L.; Vergnes, L.; Gozo, M.; Beach, J.A.; Salumbides, B.; Reue, K.; Wiedemeyer, W.R.; Nielsen, J.; et al. Succinate dehydrogenase inhibition leads to epithelial-mesenchymal transition and reprogrammed carbon metabolism. Cancer Metab. 2014, 2, 1–15. [Google Scholar] [CrossRef]
- Aarnoutse, R.; Ziemons, J.; Penders, J.; Rensen, S.S.; De Vos-Geelen, J.; Smidt, M.L. The clinical link between human intestinal microbiota and systemic cancer therapy. Int. J. Mol. Sci. 2019, 20, 4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vétizou, M.; Daillère, R.; Zitvogel, L. Microbiote intestinal et réponses aux thérapies anti-tumorales. Med. Sci. 2016, 32, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Dzutsev, A.; Goldszmid, R.S.; Viaud, S.; Zitvogel, L.; Trinchieri, G. The role of the microbiota in inflammation, carcinogenesis, and cancer therapy. Eur. J. Immunol. 2015, 45, 17–31. [Google Scholar] [CrossRef]
- Spanogiannopoulos, P.; Bess, E.N.; Carmody, R.N.; Turnbaugh, P.J. The microbial pharmacists within us: A metagenomic view of xenobiotic metabolism. Nat. Rev. Microbiol. 2016, 14, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Sougiannis, A.T.; VanderVeen, B.N.; Enos, R.T.; Velazquez, K.T.; Bader, J.E.; Carson, M.; Chatzistamou, I.; Walla, M.; Pena, M.M.; Kubinak, J.L.; et al. Impact of 5 fluorouracil chemotherapy on gut inflammation, functional parameters, and gut microbiota. Brain Behav. Immun. 2019, 80, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Wallace, B.D.; Wang, H.; Lane, K.T.; Scott, J.E.; Orans, J.; Koo, J.S.; Venkatesh, M.; Jobin, C.; Yeh, L.-A.; Mani, S.; et al. Alleviating Cancer Drug Toxicity by Inhibiting a Bacterial Enzyme. Science 2010, 330, 831–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, I.D.; Nicholson, J.K. Gut Microbiome Interactions with Drug Metabolism, Efficacy and Toxicity Europe PMC Funders Author Manuscripts The gut microbiota have the capability of preforming a wide range of metabolic reactions on. Transl. Res. 2017, 179, 204–222. [Google Scholar] [CrossRef] [Green Version]
- Haiser, H.J.; Turnbaugh, P.J. Metagenomic View of Xenobiotic Metabolism. Pharmacol. Res. 2013, 69, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringer, A.M.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Yeoh, A.S.J.; Keefe, D.M.K. Faecal microflora and β-glucuronidase expression are altered in an irinotecan-induced diarrhoea model in rats. Cancer Biol. Ther. 2008, 7, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, F.M.; Maison, N.; Holtrop, G.; Young, P.; Stevens, V.J.; Ince, J.; Johnstone, A.M.; Lobley, J.E.; Flint, H.J.; Louis, P. Phylogenetic distribution of genes encoding β-glucuronidase activity in human colonic bacteria and the impact of diet on faecal glycosidase activities. Environ. Microbiol. 2012, 14, 1876–1887. [Google Scholar] [CrossRef]
- Carmody, R.N.; Turnbaugh, P.J. Host-microbial interactions in the metabolism of therapeutic and diet-derived xenobiotics. J. Clin. Investig. 2014, 124, 4173–4181. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharlau, D.; Borowicki, A.; Habermann, N.; Hofmann, T.; Klenow, S.; Miene, C.; Munjal, U.; Stein, K.; Glei, M. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res. Rev Mutat. Res. 2009, 682, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Leonel, A.J.; Alvarez-Leite, J.I. Butyrate: Implications for intestinal function. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 474–479. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Maria, R.M.; Altei, W.F.; Selistre-de-Araujo, H.S.; Colnago, L.A. Impact of chemotherapy on metabolic reprogramming: Characterization of the metabolic profile of breast cancer MDA-MB-231 cells using 1H HR-MAS NMR spectroscopy. J. Pharm. Biomed. Anal. 2017, 146, 324–328. [Google Scholar] [CrossRef] [Green Version]
- Schug, Z.T.; VandeVoorde, J.; Gottlieb, E. The metabolic fate of acetate in cancer. Nat. Rev. Cancer 2016, 16, 708–717. [Google Scholar] [CrossRef]
- Salonen, A.; Lahti, L.; Salojärvi, J.; Holtrop, G.; Korpela, K.; Duncan, S.H.; Date, P.; Farquharson, F.; Johnstone, A.M.; Lobley, G.E.; et al. Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 2014, 8, 2218–2230. [Google Scholar] [CrossRef]
- Lamichhane, S.; Yde, C.C.; Schmedes, M.S.; Jensen, H.M.; Meier, S.; Bertram, H.C. Strategy for Nuclear-Magnetic-Resonance-Based Metabolomics of Human Feces. Anal. Chem. 2015, 87, 5930–5937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Cui, Q.; Lewis, I.A.; Hegeman, A.D.; Anderson, M.E.; Li, J.; Schulte, C.F.; Westler, W.M.; Eghbalnia, H.R.; Sussman, M.R.; Markley, J.L. Metabolite identification via the Madison Metabolomics Consortium Database. Nat. Biotechnol. 2008, 26, 162–164. [Google Scholar] [PubMed]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0-The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, 801–807. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | |
|---|---|
| Number | 8 |
| Sex (M/F) | F |
| Age | 62.4 ± 4 |
| BMI (Kg/m2) | 28.04 ± 5 |
| Infected Breast | 2 L/5 R/1 both |
| Histology | IDC |
| Receptors | ER+, PR+ |
| Grade | SBR II |
| Treatment | FEC |
| Treatment response | Six good-responders and two non-responders |
| Diet | Balanced diet |
| Physical activities | No physical activities |
| Collect time | Early morning |
| Compound | 1H Chemical Shift (ppm) | 13C Chemical Shift (ppm) |
|---|---|---|
| Acetate | 1.9204(s) | 26.09 |
| Acetoacetate | 2.262 (s); 3.421 (s) | 32.254; 56.1864 |
| Acetone | 2.299 (s) | 33.4 |
| Alanine | 1.477 (d); 3.789 (q) | 15.99; 21.86; 40.45 |
| Alpha-Glucose | 3.406; 3.531; 5.238 (d) | 72.352; 74.166; 94.826 |
| Alpha-Xylose | 5.209 (d) | 94.959 |
| Arabinose | 4.524 (d); 5.271 (d) | 99.61; 96.14 |
| Aspartate | 2.684 (dd); 2.817(dd); 3.904 (dd) | 39.23; 54.92; 177.04 |
| Anserine | 3.772 (s) | 35.16 |
| Beta-Glucose | 3.249 (dd); 3.494; 4.652 (d) | 76.854; 78.690; 98.643 |
| Beta-Xylose | 4.582 (d); 3.318 | 99.352; 76.753 |
| Betaine | 3.255 (s); 3.886 (s) | 56.019; 68.839 |
| Butyrate | 0.898 (t); 1.561 (q); 2.16 (t) | 15.99; 21.86; 40.45 |
| Caffeine | 3.353 (s); 3.511 (s); 3.947 (s) | 30.609; 32.517; 35.967 |
| Choline | 3.199 (s) | 56.618 |
| Creatine | 3.034 (s); 3.937 (s) | 39.653; 56.535 |
| Creatinine | 3.046 (s); 4.06 (s) | 32.936; 59 |
| Desaminotyrosine | 2.448 (t); 6.854 (d) | 29.254; 115.4315 |
| 3.4-Dihydroxybenzeneacetate | 3.382 (s); 6.692 (dd); 6.778 (d) | 46.169; 124.12; 119.553 |
| 1.3-Dihydroxyacetone | 4.413 (s) | 67.535 |
| 1.7-Dimethylxanthine | 3.305 (s); 3.926 (s) | 30.375; 35.839 |
| Ethanol | 1.187 (t); 3.659 (q) | 19.58; 60.167 |
| Ferulic acid | 6.899 (d) | 117.8 |
| Formate | 8.46 (s) | |
| Fumarate | 6.522 (s) | 138.1 |
| D-Galactose | 4.084 (t); 5.262 (d) | 74.591; 95.014 |
| Glutamate | 3.763 (m) | 57.357 |
| Guanosine | 7.985 (s) | 140.4 |
| Glycerol | 3.657 (m) | 65.187 |
| Glycine | 3.564 (s) | 44.308 |
| Glycolate | 3.930 (s) | 63.929 |
| Histamine | 7.144 (s); 7.913(s) | 119.02; 138.6 |
| Histidine | 7.085 (s); 7.852 (s) | 119.6; 138.8 |
| Homovanillate | 3.439 (s); 3.85 (s); 6.756 (dd) | 46.586; 58.594; 124.638 |
| 2-Hydroxyisovalerate | 0.836 (d); 3.844 (d) | 18.26; 79.98 |
| 3-Hydroxyisovalerate | 2.354 (s) | 2.354 (s) |
| Hypoxanthine | 7.982 (s); 8.116 (s) | 144.43; 140.08 |
| Indole-3-acetate | 3.645 (s); 7.617 (d) | 36.548; 121.38 |
| Isobutyrate | 1.065 (d); 2.39 (m) | 22.102; 39.672 |
| Isoleucine | 0.906 (t); 1.003 (d); 3.655 (d) | 13.834; 17.411; 62.249 |
| Isopropyl alcohol | 1.177 (d); 4.024 (m) | 26.325; 67.064 |
| Isovalerate | 0.904 (d); 2.058 (d) | 24.67; 49.906 |
| Kynurenine | 3.707 (d) | 41.678 |
| Lactate | 1.312 (d); 4.107 (q) | 23.812; 72.127 |
| Levulinic acid | 2.39 (t) | 34.012 |
| Malonic acid | 3.113 (s) | 50.242 |
| Maltose | 5.21 (d); 5.387 (d) | 94.613; 102.274 |
| Methanol | 3.36 (s) | 51.571 |
| Methionine | 2.141 (s) | 32.721 |
| Methylamine | 2.606 (s) | 27.603 |
| 3-Methylhistidine | 3.721 (s); 7.065 (s) | 34.49; 127.022 |
| Myo-Inositol | 4.104 (t); 3.27 | 74.93; 77.16 |
| N-Acetylglycine | 2.029 (s) | 22.322 |
| N.N-Dimethylglycine | 2.918(s); 3.709 (s) | 46.229; 62.578 |
| Nicotinate | 8.942 (s); 8.253; 8.613 | |
| Ornithine | 3.034 (t) | 42.2 |
| Orotic acid | 6.199 (s) | 104.059 |
| p-Cresol | 2.247 (s) | 22.0728 |
| Phenylacetate | 3.54(s); 7.304 (m); 7.374 (m) | 47.174; 129.162; 131.868 |
| Phenylalanine | 7.336 (m); 7.374 (m); 7.434 (m) | 132.09; 129.1; 131.8 |
| Hydrocinnamic acid | 2.495 (t); 2.898 (t) | 41.873; 34.561 |
| Propionate | 1.059 (t); 2.184 (m) | 12.872; 33.4 |
| Pyruvate | 2.366 (s) | 29.21 |
| Ribose | 4.106; 4.217 (dd); 4.936 (d); 5.254 (d) | 71.721; 73.411; 96.527; 99.032 |
| S-Adenosylhomocysteine | 8.373 (s) | 139.752 |
| Sarcosine | 3.598 (s) | 53.513 |
| Succinate | 2.423 (s) | 36.915 |
| Sucrose | 5.39 (d) | 94.899 |
| Syringate | 3.905 (s) | 59.06 |
| Theophylline | 3.31 (s); 3.48 (s) | 30.93; 32.95 |
| Threonine | 1.334 (d); 3.592 (d); 4.26 (m) | 22.189; 63.272; 68.779 |
| Trimethylamine N-oxide | 3.249 (s) | 62.259 |
| Tyramine | 2.933 (t); 6.908 (d); 7.223 (d) | 34.654; 118.542; 133.054 |
| Tyrosine | 6.904 (d); 7.199 (d) | 118.8; 133.3 |
| Tryptophan | 7.53 (d); 7.311 (m) | 114.5; 128 |
| U1 (Unknown) | 1.813 (s) | 25.922 |
| Uracil | 5.808 (d); 7.54 (d) | 103.794; 146.5 |
| Urocanate | 6.397 (d); 7.273 (d); 7.786 (s) | 124.4; 133.7; 140.4 |
| Valerate | 0.860 (t); 1.306 (m); 2.191 (m) | 15.75; 24.548; 40.4 |
| Valine | 0.994 (d); 1.048(d); 3.616 (d) | 19.365; 20.698; 63.082 |
| Vanillate | 3.898 (s) | 55.895 |
| Xanthine | 7.947 (s) | 141.4 |
| Metabolites | VIP | C0-C2 | C2-C3 | ROC Analysis | ||||
|---|---|---|---|---|---|---|---|---|
| FDR | FC | FDR | FC | a AUC | b Sensitivity | c Specificity | ||
| Acetate | 3.26 | <0.001 | 3.9871 | <0.001 | 4.049 | 1.0 | 1.0 | 1.0 |
| Succinate | 9.02 | <0.001 | 0.003884 | <0.001 | 0.30446 | 1.0 | 1.0 | 1.0 |
| Lactate | 2.90 | <0.001 | 0.021075 | <0.001 | 12.43 | 0.98438 | 0.938 | 1.0 |
| Glycine | 1.19 | <0.001 | 0.14787 | <0.001 | 3.5881 | 0.969 | 0.812 | 1.0 |
| Ribose | 2.16 | <0.001 | 10.817 | <0.001 | 0.015883 | 0.90625 | 0.695 | 1.0 |
| Valerate | 1.61 | <0.001 | 16.569 | <0.001 | 0.29447 | 0.89062 | 0.772 | 1.0 |
| Alanine | 2.55 | <0.001 | 0.37297 | <0.001 | 6.8116 | 0.89062 | 0.836 | 1.0 |
| Valine | 1.53 | <0.001 | 120.41 | <0.001 | 0.35782 | 0.875 | 0.699 | 1.0 |
| Phenylacetate | 1.63 | <0.001 | 2.4325 | <0.001 | 0.53583 | 0.85938 | 0.688 | 1.0 |
| Ethanol | 1.86 | <0.001 | 0.37961 | <0.001 | 2.1783 | 0.85938 | 0.671 | 1.0 |
| Butyrate | 2.39 | <0.001 | 3.3069 | <0.001 | 0.39693 | 0.84375 | 0.641 | 1.0 |
| Isoleucine | 2.64 | <0.001 | 0.0039568 | <0.001 | 0.22841 | 0.82031 | 0.547 | 1.0 |
| Creatinine | 1.99 | <0.001 | 10.631 | <0.001 | 0.16213 | 0.82031 | 0.751 | 1.0 |
| Isobutyrate | 3.15 | <0.001 | 2259.1 | <0.001 | 0.075646 | 0.8125 | 0.625 | 1.0 |
| Arabinose | 3.31 | <0.001 | 87.253 | <0.001 | 0.015883 | 0.75 | 0.762 | 0.936 |
| Threonine | 1.33 | <0.001 | 2.1585 | <0.001 | 2.2738 | 0.719 | 0.344 | 1.0 |
| Xylose | 2.83 | <0.001 | 89.196 | <0.001 | 1.9863 | 0.71875 | 0.638 | 0.938 |
| Propionate | 2.32 | <0.001 | 4.7322 | <0.001 | 0.6252 | 0.703 | 0.638 | 0.953 |
| Glutamate | 1.07 | <0.001 | 0.0031898 | <0.001 | 0.23818 | 0.6875 | 0.662 | 0.875 |
| 3.4-Dihydroxybenzeneacetate | 1.35 | <0.001 | 0.0099035 | <0.001 | 838.3 | 0.6875 | 0.653 | 0.836 |
| Isovalerate | 1.78 | <0.001 | 0.2328 | <0.001 | 0.44677 | 0.63281 | 0.632 | 0.895 |
| Aspartate | 1.59 | <0.001 | 0.089599 | <0.001 | 0.5052 | 0.625 | 0.65 | 0.812 |
| Ornithine | 1.48 | <0.001 | 0.17305 | <0.001 | 83.958 | 0.625 | 0.536 | 0.844 |
| Methanol | 2.86 | <0.001 | 0.1776 | <0.001 | 3.8184 | 0.61719 | 0.344 | 0.863 |
| Fumarate | 1.73 | <0.001 | 12.937 | <0.001 | 3.2866 | 0.60938 | 0.635 | 0.961 |
| N.N-Dimethylglycine | 1.77 | <0.001 | 0.032145 | <0.001 | 7.3135 | 0.58594 | 0.634 | 0.875 |
| Theophylline | 1.46 | <0.001 | 0.41299 | <0.001 | 0.09096 | 0.52344 | 0.625 | 0.766 |
| Compounds | GR p-Value t-Test after vs. before Chemotherapy | PR p-Value t-Test after vs. before Chemotherapy | Compounds | GR p-Value t-Test after vs. before Chemotherapy | PR p-Value t-Test after vs. before Chemotherapy |
|---|---|---|---|---|---|
| Alanine | 0.002 ↑ | 0.0869 | Lactate | 0.1021 | 0.1166 |
| Succinate | 0.0016 ↑ | 0.1734 | 1.3-Dihydroxyacetone | 0.1033 | 0.2504 |
| Glutamate | 0.0042 ↑ | 1 | Arabinose | 0.1049 | 0.25 |
| Tyrosine | 0.1612 | 0.0059 ↑ | Betaine | 0.1266 | 0.25 |
| Fumarate | 0.0061 ↓ | 0.1907 | Valerate | 0.1274 | 0.25 |
| 3-Methylhistidine | 0.0063 ↑ | 0.0085 ↓ | Alpha-Xylose | 0.1319 | 0.2102 |
| Acetate | 0.0079 ↑ | 0.1554 | Vanillate | 0.1328 | 1 |
| Propionate | 0.0081 ↑ | 0.1063 | 1.7-Dimethylxanthine | 0.1415 | 1 |
| Creatine | 0.0085 ↑ | 0.1248 | D-Galactose | 0.1513 | 0.25 |
| Hypoxanthine | 0.0087 ↓ | 0.2471 | Methylamine | 0.1514 | 0.25 |
| Histamine | 0.0105 ↑ | 0.2404 | Beta-Xylose | 0.1554 | 0.1920 |
| Valine | 0.0120 ↑ | 0.2015 | Beta-Glucose | 0.1570 | 0.2840 |
| Methionine | 0.0148 ↑ | 0.0506 | Formate | 0.1587 | 0.2533 |
| Ethanol | 0.0157 ↑ | 0.25 | Isobutyrate | 0.1623 | 0.2534 |
| Glycine | 0.0160 ↑ | 0.2299 | Alpha-Glucose | 0.1663 | 0.2796 |
| Butyrate | 0.0514 | 0.0165 ↓ | N.N-Dimethylglycine | 0.1663 | 0.2601 |
| Isoleucine | 0.0201 ↑ | 0.1476 | Levulinic acid | 0.1746 | 0.7987 |
| Phenylacetate | 0.0236 ↑ | 0.2791 | Choline | 0.17854 | 0.2934 |
| U1 (Unknown) | 0.0322 | 0.25 | Anserine | 0.1816 | 0.25 |
| Methanol | 0.1542 | 0.0343 ↓ | Isopropanol | 0.1816 | 0.25 |
| Uracil | 0.0885 | 0.0348 ↑ | Tyramine | 0.181609 | 1 |
| Acetone | 0.0831 | 0.0365 ↑ | Trimethylamine N-oxide | 0.1816 | 1 |
| Theophylline | 0.0379 ↑ | 0.1693 | Orotic acid | 0.1816 | 1 |
| Isovalerate | 0.0560 | 0.3306 | Maltose | 0.1816 | 1 |
| Ribose | 0.0602 | 0.4583 | 3-Hydroxyisovalerate | 0.2066 | 0.25 |
| Caffeine | 0.0619 | 0.2692 | Glycolate | 0.2383 | 0.25 |
| Sarcosine | 0.0651 | 0.0832 | Desaminotyrosine | 0.2466 | 0.25 |
| Homovanillate | 0.0667 | 0.25 | Myo-Inositol | 0.2704 | 1 |
| Sucrose | 0.0778 | 0.7901 | Ferulic acid | 0.2793 | 1 |
| Pyruvate | 0.2468 | 0.0847 | Xanthine | 0.2866 | 0.2113 |
| Creatinine | 0.0864 | 0.1824 | Tryptophan | 0.2849 | 0.2044 |
| Aspartate | 0.0873 | 1 | Acetoacetate | 0.3167 | 0.1917 |
| N-Acetylglycine | 0.0873 | 0.25 | Urocanate | 0.3255 | 0.1091 |
| p-Cresol | 0.0873 | 0.2596 | S-Adenosylhomocysteine | 0.3265 | 0.25 |
| Lysine | 0.0873 | 1 | Ornithine | 0.3321 | 0.2592 |
| Malonic acid | 0.0882 | 0.2803 | 3,4-Dihydroxybenzeneacetate | 0.3632 | 1 |
| Phenylalanine | 0.3103 | 0.0964 | Syringate | 0.3837 | 0.3645 |
| Glycerol | 0.0991 | 0.2764 | Nicotinate | 0.4037 | 0.3360 |
| Threonine | 0.1318 | 0.0990 | Histidine | 0.4625 | 0.25 |
| Proline | 0.0996092 | 0.25 | Guanosine | 0.5633 | 0.1791 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zidi, O.; Souai, N.; Raies, H.; Ben Ayed, F.; Mezlini, A.; Mezrioui, S.; Tranchida, F.; Sabatier, J.-M.; Mosbah, A.; Cherif, A.; et al. Fecal Metabolic Profiling of Breast Cancer Patients during Neoadjuvant Chemotherapy Reveals Potential Biomarkers. Molecules 2021, 26, 2266. https://doi.org/10.3390/molecules26082266
Zidi O, Souai N, Raies H, Ben Ayed F, Mezlini A, Mezrioui S, Tranchida F, Sabatier J-M, Mosbah A, Cherif A, et al. Fecal Metabolic Profiling of Breast Cancer Patients during Neoadjuvant Chemotherapy Reveals Potential Biomarkers. Molecules. 2021; 26(8):2266. https://doi.org/10.3390/molecules26082266
Chicago/Turabian StyleZidi, Oumaima, Nessrine Souai, Henda Raies, Farhat Ben Ayed, Amel Mezlini, Sonia Mezrioui, Fabrice Tranchida, Jean-Marc Sabatier, Amor Mosbah, Ameur Cherif, and et al. 2021. "Fecal Metabolic Profiling of Breast Cancer Patients during Neoadjuvant Chemotherapy Reveals Potential Biomarkers" Molecules 26, no. 8: 2266. https://doi.org/10.3390/molecules26082266