1. Introduction
Cutaneous melanoma is a complex disorder characterized by elevated heterogeneity. It is one of the most aggressive types of cancer and one of the leading causes of skin cancer-related mortality, due to its metastatic power. Its therapeutic management is a real challenge, as it is amongst the solid malignancies most refractory to conventional cancer therapies [
1]. Up to 90% of melanomas exhibit aberrant MAPK pathway activation that induces cell cycle deregulation and apoptosis inhibition, and marked improvements in cutaneous melanoma treatment have been achieved by targeting the MAPK signaling pathway. Improved overall survival outcomes were observed with targeted therapies in patients with BRAF
V600 mutant unresectable stage III or stage IV melanoma. Nearly half of patients with metastatic melanomas harbor a valine to glutamine substitution in codon 600 of the serine/threonine kinase BRAF [
2]. Vemurafenib, dabrafenib, and encorafenib are BRAF inhibitors (BRAFi) approved by the US Food and Drug Administration (FDA) to treat patients with BRAF
V600E-mutated metastatic melanomas [
3]. BRAFi have relatively high response rates; however, patients almost invariably develop disease progression after about five months. The addition of a MEK inhibitor such as trametinib to BRAFi extends the median duration of response from five months to nine months [
4,
5]. However, many patients develop resistance to BRAF (+/− MEK) inhibitors [
5,
6].
Amongst the mechanisms used by cancer cells to become resistant to treatment, multidrug resistance (MDR) has been intensively studied [
7]. The most prominent mechanism underlying MDR is the overexpression of multidrug transporters, of which the best known in cancers were ATP-binding cassette (ABC) transporters [
8]. However, to date, the FDA has not approved the use of any ABC transporter inhibitor, due to toxicity issues [
9]. Therefore, counteracting chemotherapy resistance by alleviating MDR is still an unmet medical need.
We recently discovered that the Hedgehog receptor Ptch1, which is overexpressed in many cancers, has an efficient drug efflux function in cancer cells. We have demonstrated that Ptch1 pumps chemotherapeutic agents such as doxorubicin (dxr) which isused to treat many cancers, out of cancer cells that were derived from melanoma and adrenocortical carcinoma (ACC), thereby conferring resistance to chemotherapy [
10,
11,
12]. Our analysis of normalized gene expression data and matching clinical information for cutaneous melanoma tumors downloaded from The Cancer Genome Atlas (TCGA) revealed that the Ptch1 protein was strongly expressed in metastatic samples from melanoma patients whose melanoma did or did not present the BRAF
V600 mutation [
12]. In this cohort, the Kaplan–Meier analysis for a subset of patients with metastatic disease who did not receive immunotherapy indicated that a high level of Ptch1 in patient samples significantly correlated with a lower overall survival time.
We developed screening tests to identify molecules able to inhibit the drug efflux activity of Ptch1, which led to the discovery of three inhibitors [
13]. The first, panicein A hydroquinone (PAH), a compound purified from a marine sponge, increases the cytotoxicity of dxr and of the BRAF inhibitor vemurafenib against melanoma cells in vitro and in vivo [
12,
14]. The second inhibitor, methiothepin, a nonselective 5-HT receptor antagonist, increases the efficacy of dxr against adrenocortical carcinoma cells in vitro and in vivo [
11]. The third inhibitor, astemizole, an anti-histaminergic drug, also increases the efficacy of dxr against adrenocortical carcinoma cells in vitro [
15].
In the present study, we analyze the effect of methiothepin on melanoma cells. We show that methiothepin enhances the efficacy of doxorubicin against melanoma cells by binding Ptch1 and inhibiting Ptch1 efflux activity. We also report results demonstrating that methiothepin increases the effectiveness of the BRAF inhibitor vemurafenib and of the MEK inhibitor trametinib against resistant BRAFV600E melanoma cells.
3. Discussion
Although significant progress has been made in therapeutic approaches, cutaneous melanoma is still a major problem worldwide, due to its high incidence and the lack of a curative treatment for advanced stages. The discovery of new molecules for treating advanced melanomas that are resistant to existing therapies is paramount to further improve patient outcomes.
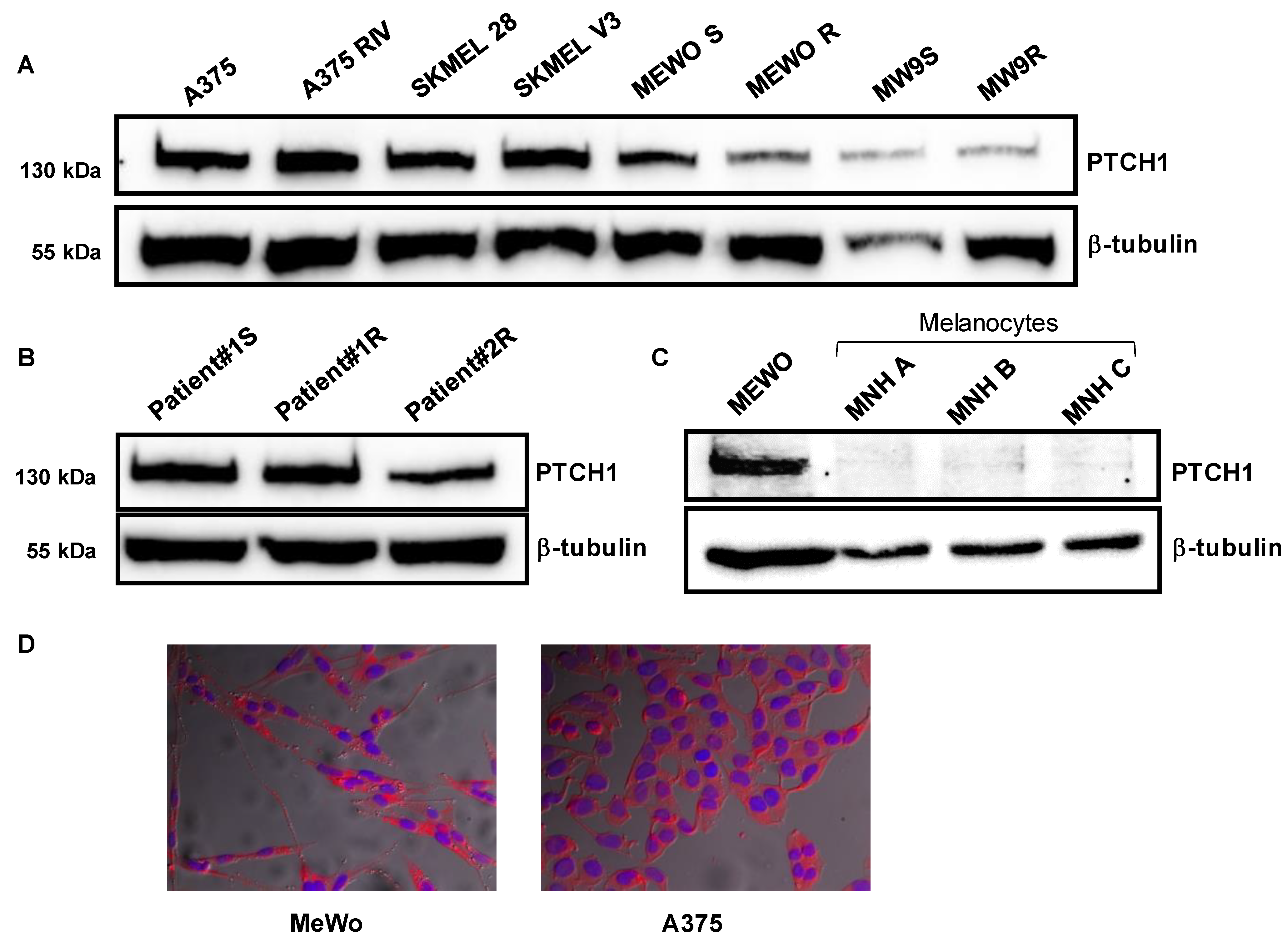
We recently reported that the Hedgehog receptor Ptch1 is strongly expressed in metastatic samples from a cohort of melanoma patients, and that a high expression level of Ptch1 in patient samples significantly correlated with a lower overall survival time [
12]. Accordingly, we observed that Ptch1 is endogenously expressed in various melanoma cell lines carrying or not a BRAF mutation. We previously found that decreased Ptch1 expression in MeWo and A375 melanoma cells using silencing RNA against Ptch1 strongly inhibited the efflux of doxorubicin, indicating that Ptch1 is involved in doxorubicin efflux in melanoma cells carrying or not the BRAF mutation [
12]. These observations allowed us to propose Ptch1 as a new target to fight melanoma resistant to treatment, recurrence and metastases.
We then performed a screening of several chemical libraries on yeast-expressing human Ptch1, looking for the inhibition of the resistance to doxorubicin conferred by Ptch1 to yeast, and identified three molecules (panicein A hydroquinone, methiothepin, and astemizole). We showed that these molecules specifically inhibited the efflux of doxorubicin from yeast-expressing Ptch1 [
11,
14,
15]. We recently provided evidence that panicein A hydroquinone not only increased the effect of classical chemotherapeutic treatments such as doxorubicin and cisplatin but also enhanced the effect of the BRAF inhibitor vemurafenib against BRAF mutated melanoma cells in vitro and in vivo [
12]. Here, we report that methiothepin, which has been shown to increase the efficacy of doxorubicin against adrenocortical carcinoma [
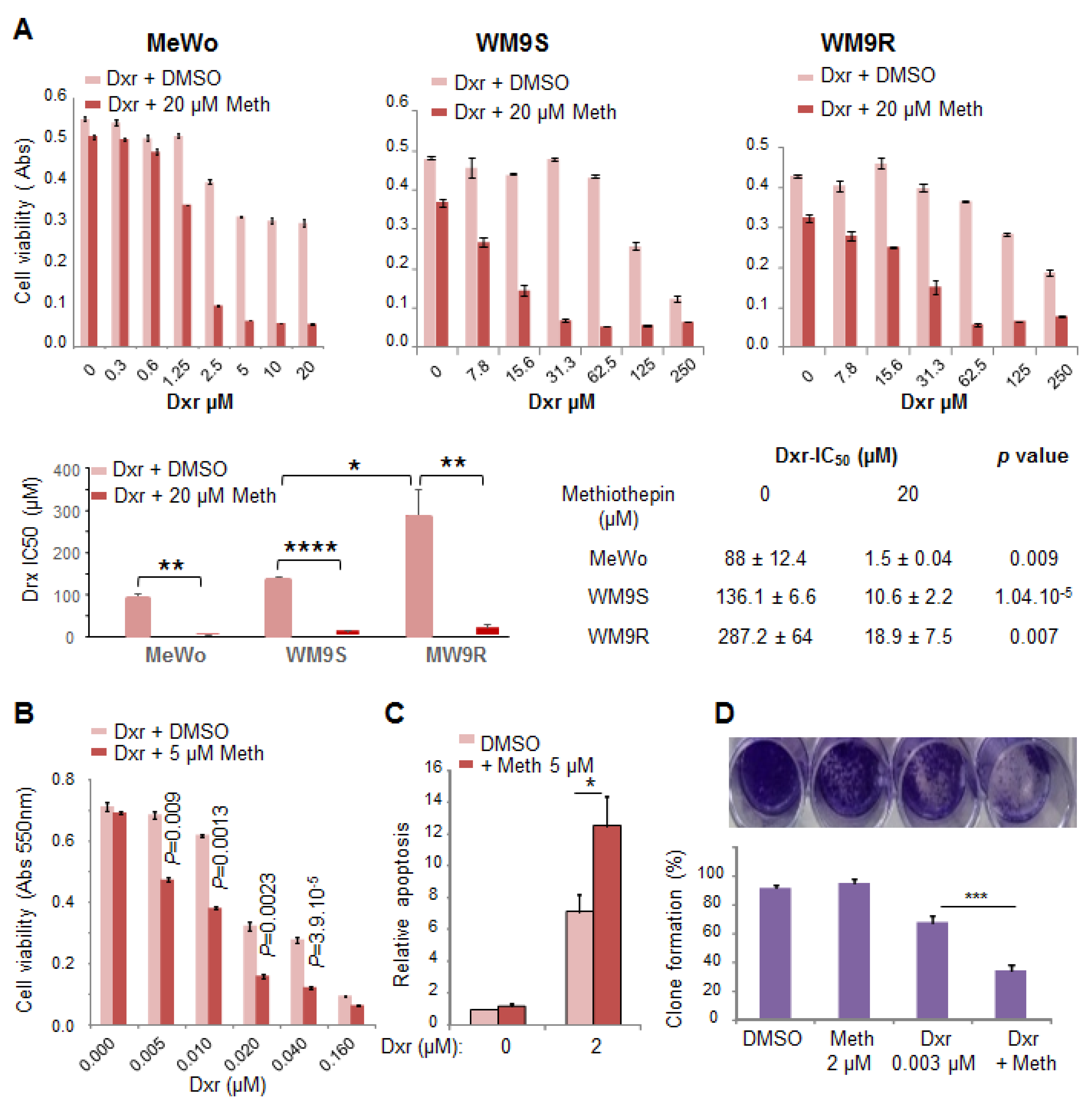
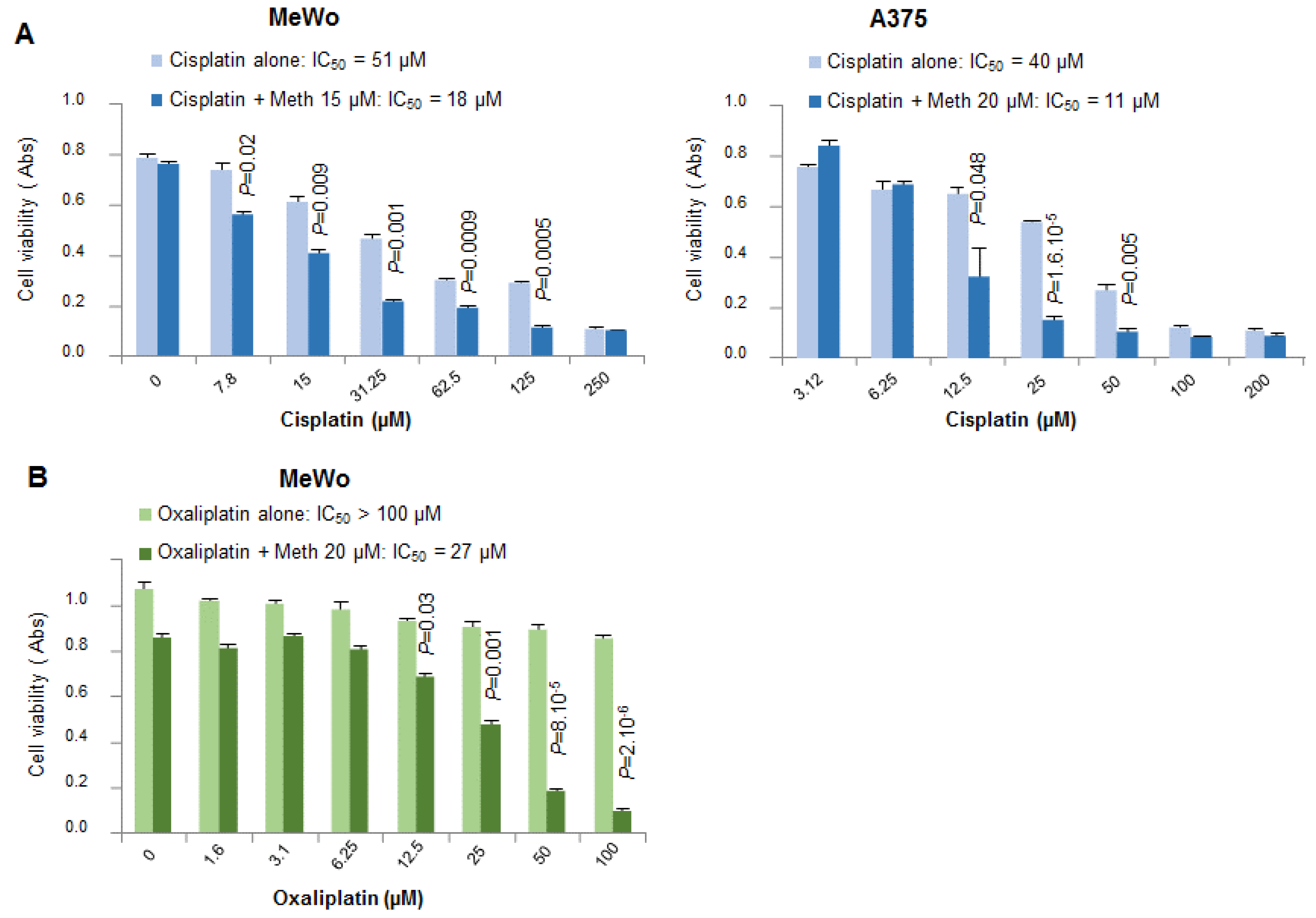
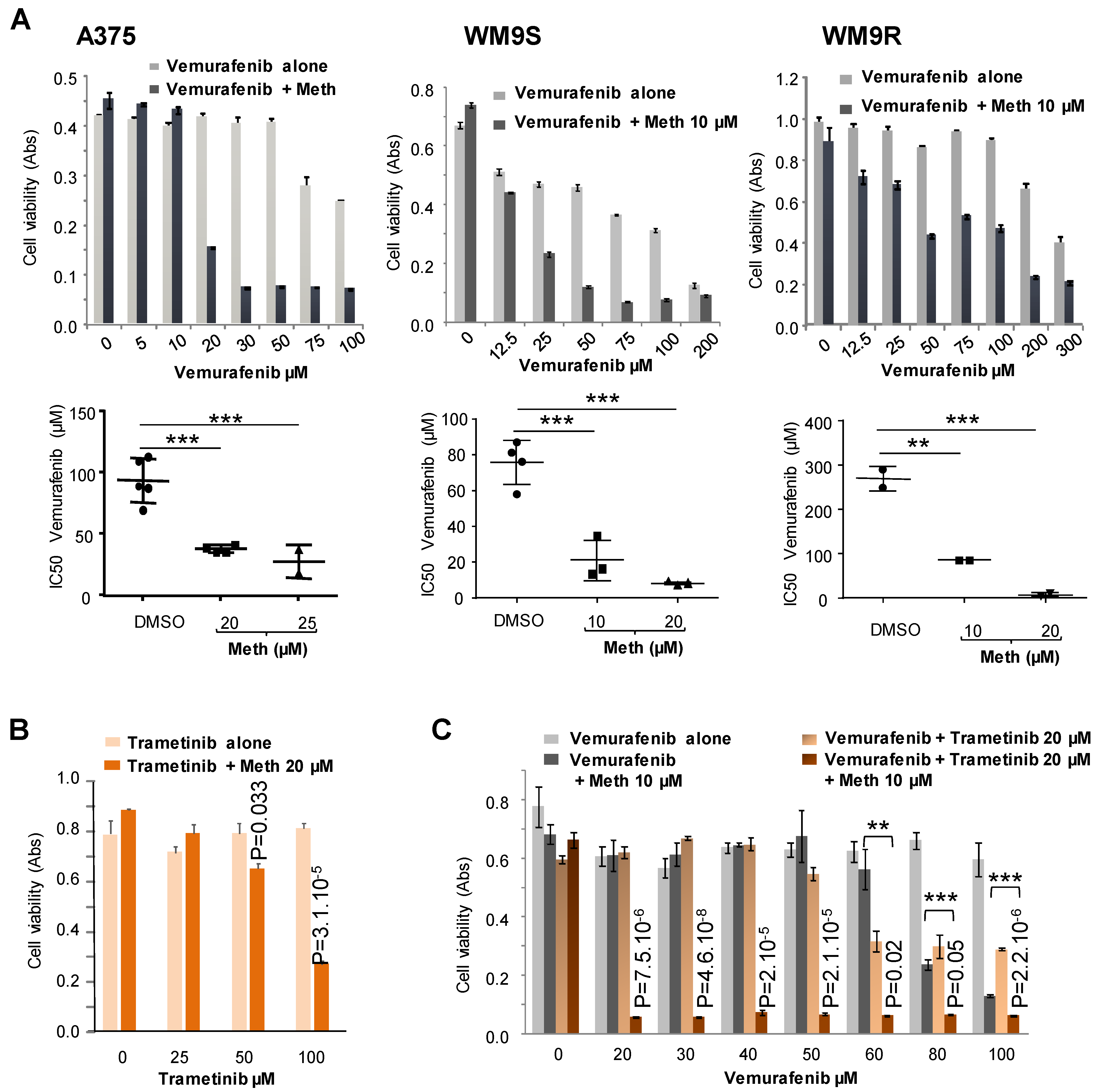
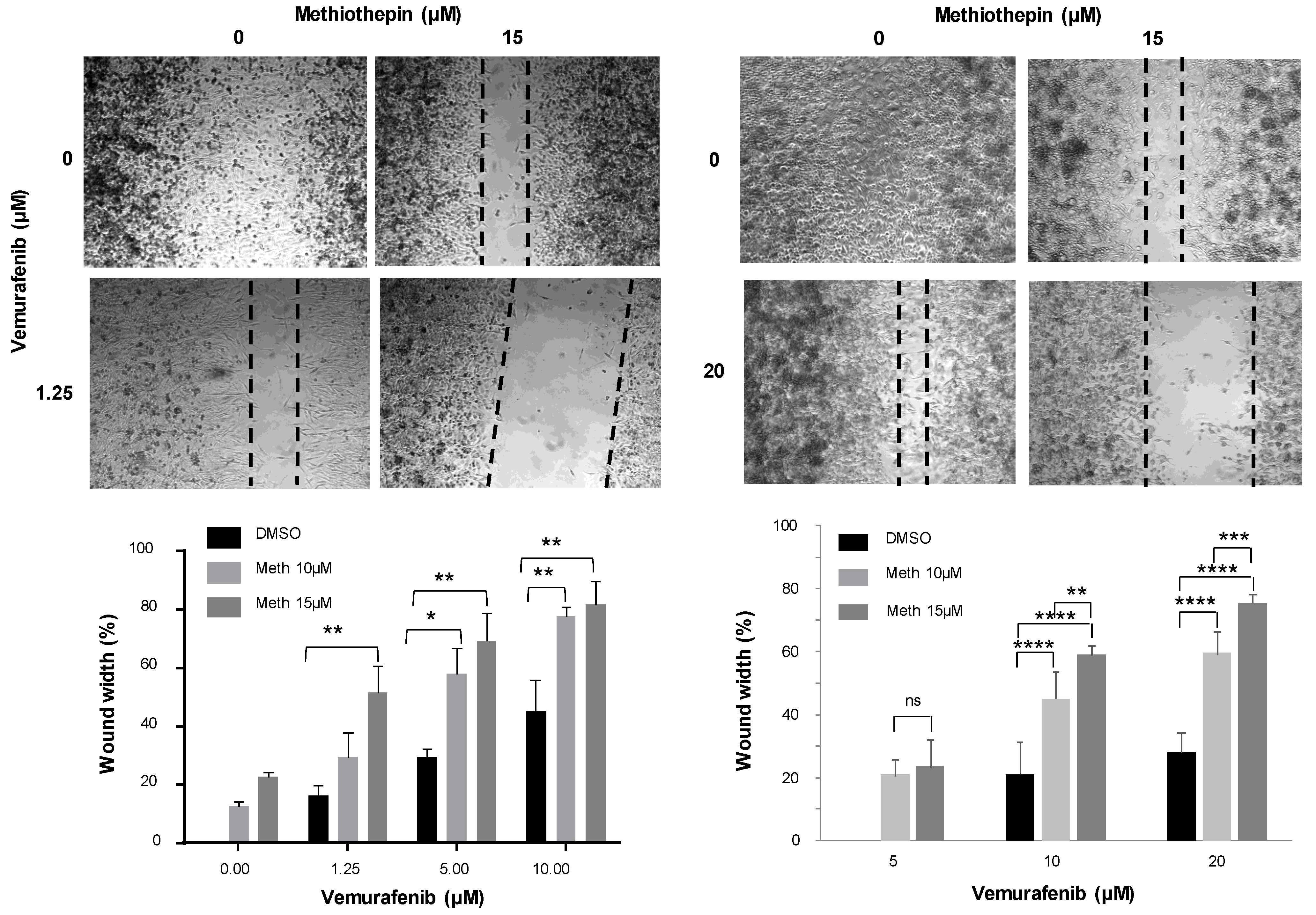
11], clearly exhibits the same effect as panicein A hydroquinone on melanoma cells. Indeed, our results demonstrate that the binding of methiothepin to Ptch1 increases the cytotoxicity of five different chemotherapeutic drugs (doxorubicin, cisplatin, oxaliplatin, vemurafenib and trametinib) against various melanoma cell lines carrying or not a BRAF mutation. We observed that the addition of methiothepin to vemurafenib prevents cell migration more efficiently than vemurafenib alone. Remarkably, these effects have also been observed on melanoma cells resistant to vemurafenib, indicating that the addition of methiothepin to vemurafenib allows vemurafenib to eliminate resistant melanoma cells and potentially the relapse of the primary tumor and the formation of metastases. Moreover, these effects were obtained at a concentration of chemotherapeutic drugs or methiothepin that did not affect by itself, suggesting that the use of methiothepin could allow to decrease the concentration of chemotherapeutic agents, and, therefore, of side effects for patients.
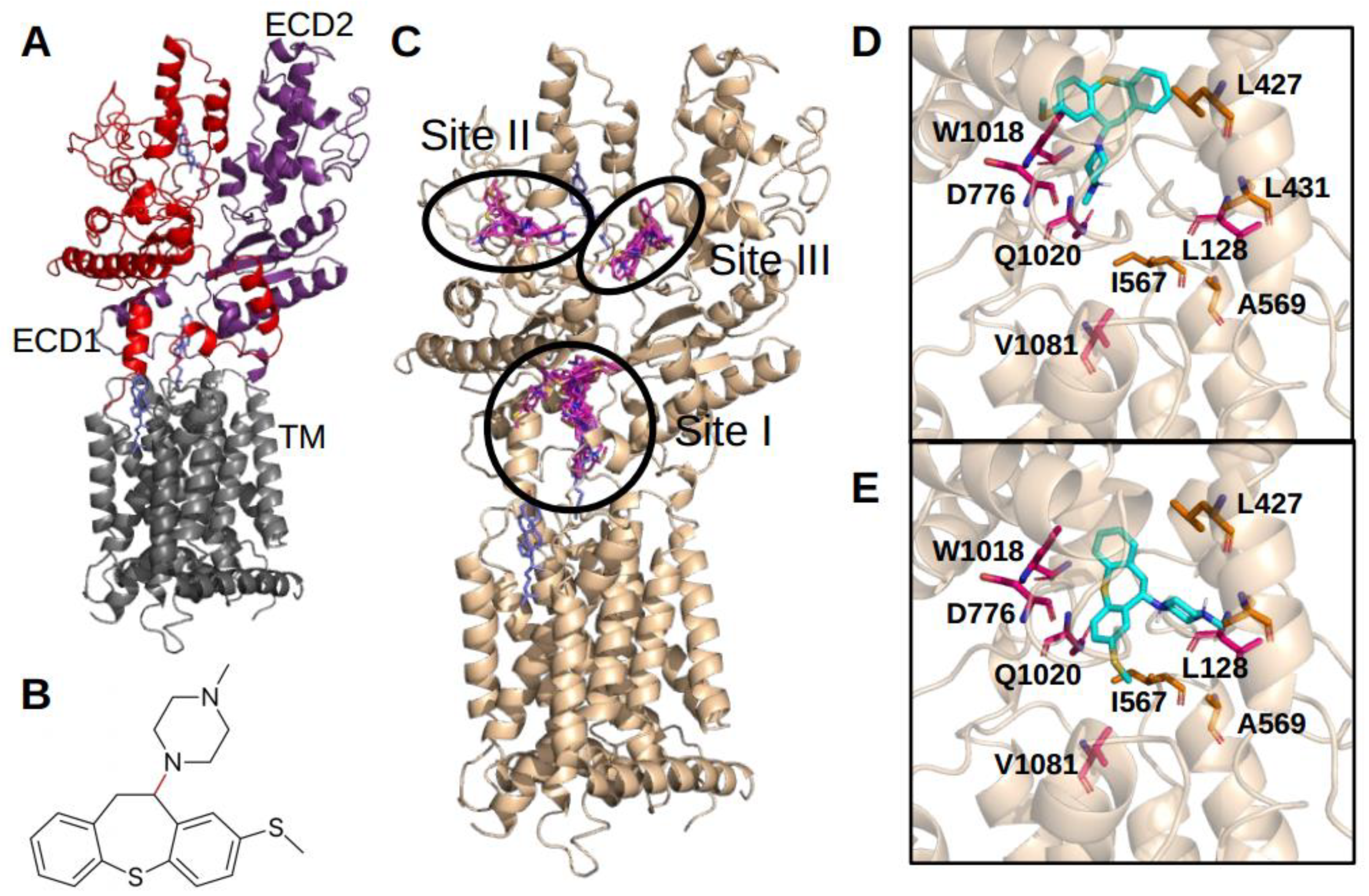
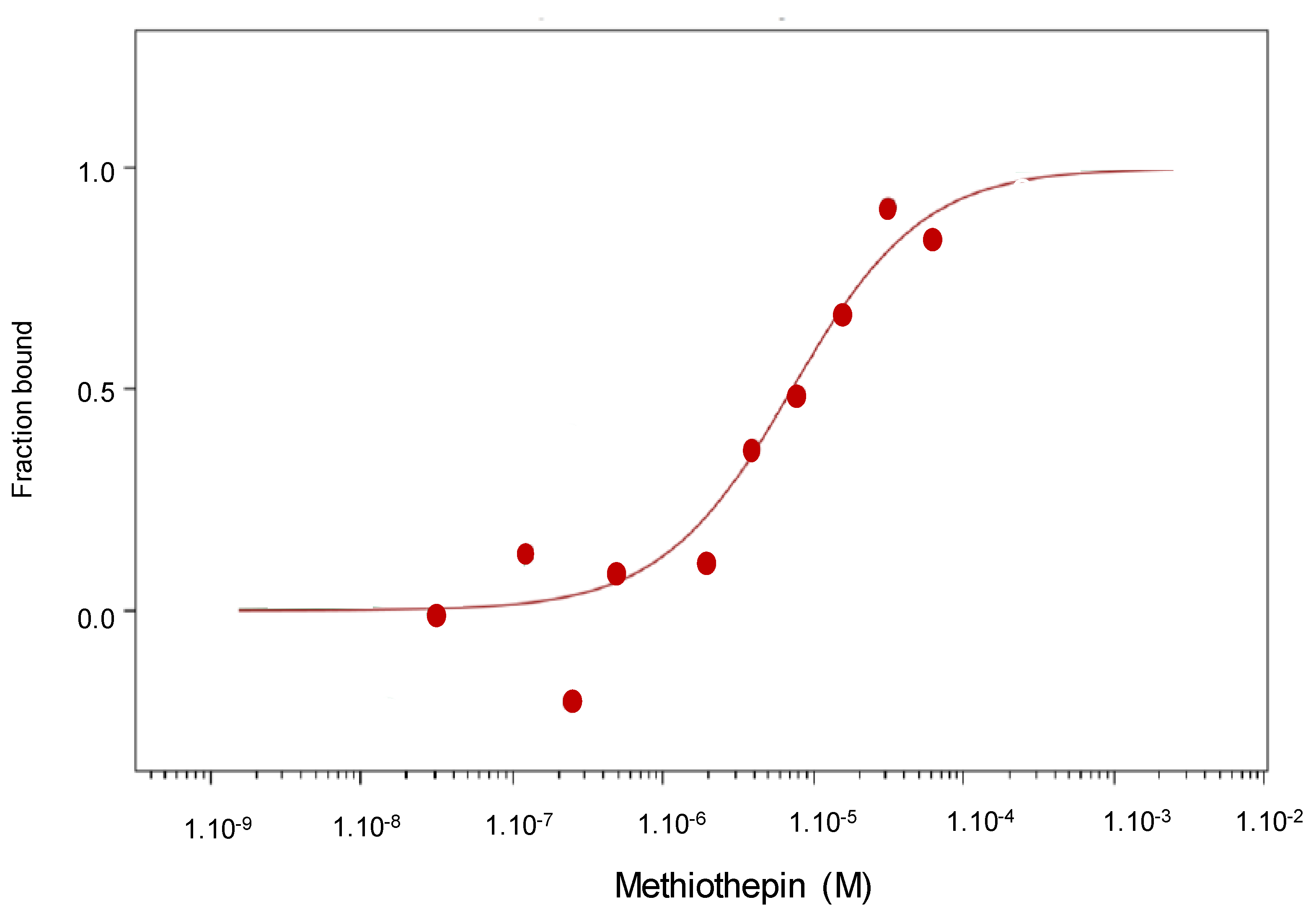
Experiments performed using microscale thermophorese technology allowed us to demonstrate that methiothepin directly interacts with Ptch1 with a Kd of 7 µM as it is the case for panicein A hydroquinone [
12]. Remarkably, our docking results suggest that methiothepin binds to Ptch1 in the central cavity previously shown to bind cholesterol, like panicein A hydroquinone, and possibly on two other sites from Ptch1 structure. Interestingly, the binding of methiothepin in site III would allow to prevent the scissor movement described previously as a possible mechanism of cholesterol efflux [
20].
Ptch1, as other efflux pumps from the RND family, uses the proton motive force to efflux drugs. Thus, Ptch1 transports drugs out of cells by interacting with protons from the extracellular medium [
10,
21]. This is only possible in tumors that have an acidic extracellular pH due to high glucose consumption of cancer cells (Warburg effect) [
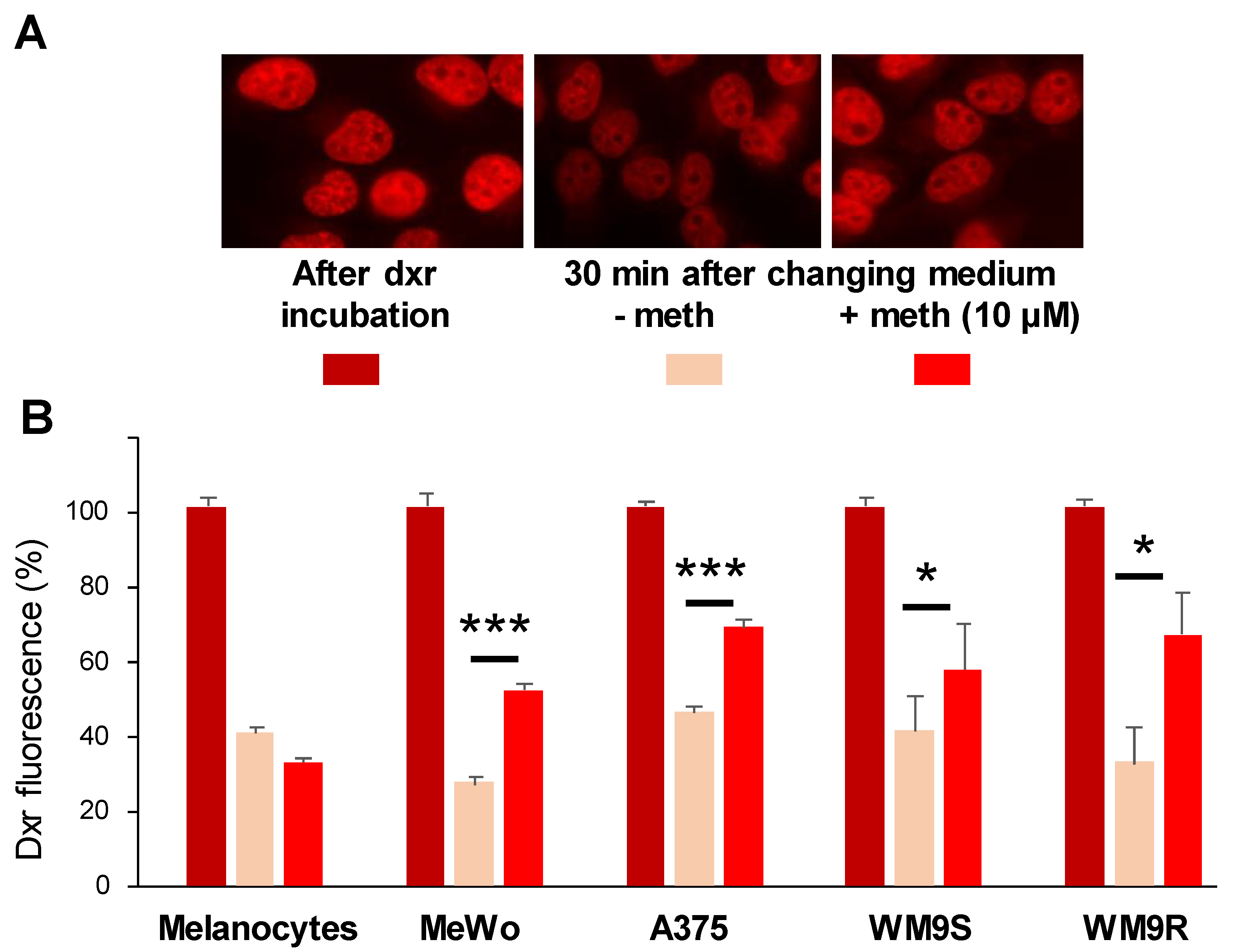
22]. This metabolic feature makes Ptch1 drug efflux activity specific to cancer cells. Consequently, Ptch1 drug efflux inhibitors can increase the concentration of chemotherapeutic agents only in cancer cells where the extracellular pH is more acidic than the intracellular one, and not in healthy cells where the extracellular medium is slightly more basic than the intracellular one preventing the formation of a proton entry gradient. A proof of the specificity of methiothepin to inhibit Ptch1 drug efflux activity only in cancer cells was brought by the quantification of doxorubicin in the tumors and the hearts of mice xenografted with adrenocortical carcinoma cells [
11]. Actually, we observed that the amount of doxorubicin in the tumors of mice treated with the combination of methiothepin and doxorubicin was significantly higher (three times) than in tumors from mice treated with doxorubicin alone, which was not the case in the heart of animals. From these results, for melanoma treatment, we can expect that methiothepin will increase chemotherapy concentration and efficacy only in melanoma cells and not in healthy tissues preventing the increase of side effects such as those observed in clinical trials with inhibitors of ABC transporters.
4. Materials and Methods
4.1. Chemical and Biological Material
Doxorubicin hydrochloride, cisplatin, and oxaliplatin were purchased from Sigma-Aldrich, St. Louis, MO, USA. Vemurafenib and Trametinib were purchased from Selleckchem. Methiothepin maleate (P375) was purchased from Santa Cruz (CAS number: 20229-30-5; MW: 472.62).
Human melanoma cell lines A375 and MeWo were purchased from ATCC. Melanocytes, other melanoma cell lines and melanoma cell cultures from patients (Patient#1S, Patient#1R and Patient#2R) were provided Robert Ballotti (C3M, Nice, France). All cells were cultured in DMEM medium supplemented with 10% fetal bovine serum and penicillin/streptomycin (Invitrogen, Villebon sur Yvette, France) at 37 °C in a 5% CO2/95% air water-saturated atmosphere.
4.2. SDS-PAGE and Western Blotting
Total RIPA extracts were prepared from melanoma cells grown to 80% confluence in 6-well plates. Protein concentrations were determined by the DC Protein Assay (Bio-Rad, Hercules, CA, USA). Samples (50 to 80 µg) were separated on SDS-PAGE and transferred to nitrocellulose membranes (Amersham, Courtaboeuf, France) using standard techniques. After 1 h at room temperature in blocking buffer (5% non-fat milk in PBS containing 0.1% Tween-20), nitrocellulose membranes were incubated overnight at 4 °C with rabbit anti-Patched antibody (Abcam ab53715; 1/1000, Cambridge, UK) or mouse anti-β-tubulin antibody (Sigma; 1/1000). After 3 washes, membranes were incubated for 45 min with anti-rabbit (1:2000) or anti-mouse (1:5000) immunoglobulin coupled to horseradish peroxidase (Dako, Courtaboeuf, France). Detection was carried out with an ECL Prime Western Blotting detection reagent (Amersham, Courtaboeuf, France) on a Fusion FX imager (Vilber Lourmat, Collegien, France), and analyses were performed using ImageJ software.
4.3. Immunofluorescence
Cells were seeded on coverslips in 24-well plates and grown to 80% confluence. Coverslips were washed twice with PBS, incubated 15 min with 4% paraformaldehyde (PFA), and then 20 min with PBS/Triton 0.1% to permeabilize cells. After blocking 30 min in PBS/2% BSA, slices were incubated at 4 °C overnight with rabbit anti-Patched antibody (Abcam ab53715; 1/1000) in PBS/0.1% BSA. After three washes in PBS/0.1% BSA, slides were incubated at room temperature during 1 h with a secondary anti-rabbit antibody coupled to Alexa 594 in PBS/0.1% BSA and washed three times before mounting using antifade reagent containing DAPI (SlowFade Gold Invitrogen, Villebon sur Yvette, France) to stain nuclei. Images were acquired with a Zeiss Axioplan 2 fluorescence microscope coupled to a digital charge-coupled device camera using a 40×/1.3 Plan NeoFluar objective and filters for Alexa 594.
4.4. Efflux Measurements
Dxr efflux measurements were carried out as previously described [
11]. Cells were seeded on coverslips in 24-well plates (Falcon, Fisher Sci. Illkirch, France) and grown to 80% confluence. Coverslips were incubated at 37 °C and 5% CO
2 with 10 μM dxr in physiological buffer (140 mM NaCl, 5 mM KCl, 1 mM CaCl
2, 1 mM MgSO
4, 5 mM glucose, 20 mM HEPES, pH 7.4). After 2 h, three coverslips were immediately fixed with 4% PFA for the dxr loading control, rapidly washed with PBS and mounted in SlowFade Gold antifade reagent with DAPI (Invitrogen, Villebon sur Yvette, France). The other coverslips (triplicate per condition) were incubated with physiological buffer supplemented with DMSO or 10 µM of methiothepin under gentle shaking at room temperature and protected from light. After 30 min, coverslips were fixed with 4% PFA, washed and mounted as described above. Images were acquired with a Zeiss Axioplan 2 fluorescence microscope coupled to a digital charge-coupled device camera using a 40×/1.3 Plan NeoFluar objective and filters for Alexa 594. Dxr fluorescence was quantified using ImageJ software. Sampling of cells was performed randomly. About 100 cells (from three wells) were scored per condition per experiment.
4.5. Molecular Docking of Methiothepin
Semi-rigid docking of methiothepin on the Ptch1 structure was performed using the Vina toolkit [
23] in USCF Chimera [
24], as previously described [
12,
15]. The same structure of Ptch1 (pdb id 6n7h chain A) and parameters for Vina were used. Possible binding sites were first assessed performing the molecular docking with a grid including the whole structure of Ptch1. After obtaining 100 poses, clusters with at least 10% of the poses (i.e., more than 10 poses) have been identified as a binding site. Then, a docking targeting specifically the binding site of interest has been performed on a grid comprising the binding site. Finally, the best poses were analyzed by assessing the distance with the key amino acids L128, L427, L431, I567, D776, W1018, Q1020, V1081.
4.6. Microscale Thermophoresis
MicroScale Thermophoresis (MST) is a biophysical technique that measures the strength of the interaction between two molecules by detecting a variation in the fluorescence signal of a fluorescently labeled target as a result of an IR-laser induced temperature change. The range of the variation in the fluorescence signal correlates with the binding of a ligand to the fluorescent target. Experiments were carried out, as previously described [
12]. Membranes from yeast-expressing human Ptch1 were solubilized with dodecyl maltoside (DDM) and incubated at 30 µg/mL with 20 nM of the fluorescent dye NT-647 2nd gen (NanoTemper Technologies, München, Germany) to label the His-tag present at the c-terminus of Ptch1 protein. The concentration of labeled solubilized fraction was kept constant, while the concentration of non-labeled ligand methiothepin was varied between 250 µM and 15 nM. The assay was performed in PBS containing 0.5% DMSO and 0.1% DDM. After a short incubation, the samples were loaded into Monolith™ NT.115 standard treated capillaries from NanoTemper Technologies and the MST analysis was performed using the NanoTemper Technologies Monolith NT.115 (LED: 30% and MST: Medium). The fluorescence within the capillary is excited and detected through the same objective. A focused InfraRed laser is used to locally heat a defined sample volume. The MST signal of fluorescent proteins changes upon binding to methiothepin resulting in different MST traces. Titration of methiothepin results in a gradual change in MST signal, which is plotted against the methiothepin concentration to yield a dose-response curve, which has been fitted to derive the methiothepin binding constant (Kd).
4.7. Cytotoxicity Assays
Cells were seeded in 96-well plates (Falcon Fisher Scientific Illkirch, France) in triplicate and grown in medium to achieve 70% to 80% confluence. Medium was then removed and replaced with 100 µL/well of complete medium containing methiothepin or DMSO as a control. After 2 h, 100 µL of complete medium containing serial dilutions of dxr, cisplatin, oxaliplatin, vemurafenib or trametinib were added. Plates were incubated at 37 °C and 5% CO2. After 24 or 48 h, cells were incubated for 3 h at 37 °C with 100 µL/well neutral red (NR) solution (50 µg/mL in medium) following the manufacturer’s protocol. Absorbance was measured in a microplate reader (Multiskan Go Microplate Spectrophotometer from Thermo Scientific, Waltham, MA, USA). IC50, defined as the concentration that resulted in a 50% decrease in the number of live cells, were calculated using GraphPad Prism 6 software.
4.8. Apoptosis Measurements
Cells were seeded at a density of 7000 cells per well in a 96-well white polystyrene plate (Falcon) in triplicate and cultured overnight at 37 °C and 5% CO2. After removal of the medium, cells were treated 48 h with medium alone or with medium containing DMSO (control), dxr alone, methiothepin alone or dxr and methiothepin together. Quantification of caspase 3/7 activity was performed using the luminescent assay CaspaseGlo 3/7 (Promega, Charbonnières-les-Bains, France) and a luminometer (Glomax 96 Microplate Luminometer (Promega, France) following the manufacturer’s protocol.
4.9. Proliferation
Cells were seeded at a density of 5000 cells per well in 96-well plates (Falcon) in triplicate and grown for 24 h at 37 °C and 5% CO2. After removal of the medium, cells were treated with medium containing serial dilutions of dxr in the presence of DMSO (control) or methiothepin.
After 7 days at 37 °C and 5% CO2, NR assay was performed for quantification of living cells. IC50 values were calculated using GraphPad Prism 6 software.
4.10. Clone Formation
Cells were seeded in 24-well plates (Falcon) at a density of 5000 cells per well in triplicate, treated with DMSO as control, methiothepin alone, dxr alone or a combination of methiothepin and dxr, and incubated at 37 °C and 5% CO2. After 7 days, 500 µL of medium containing the respective molecules was added to the wells to maintain the medium volume in wells. After 14 days, cells were fixed with 4% PFA and incubated with 0.4% crystal violet. After 1 h, cells were washed quickly with PBS and pictures of each well were taken. Cells were then solubilized with 1% SDS and absorbance was read in a microplate reader at 550 nm (Multiskan Go Microplate Spectrophotometer, Thermo Scientific).
4.11. Wound-Healing Assay
Once cells were confluent in 24-well plates, a wound was performed with a p200 tip. Wells were washed with PBS to remove cells in suspension and filled with medium containing DMSO (control), vemurafenib alone, methiothepin alone or vemurafenib and methiothepin. Two pictures were taken at two different points of each well immediately after wounding, and 48 h after wounding with Leica DM IRB (5×). The width of the wound was measured using ImageJ software and reported as percentage final wound width/initial wound width.
4.12. Statistical Analysis
All results represent at least three independent replications. Data are shown as the mean value ± SEM. Prism 6 (GraphPad) was used to determine IC50 values and other statistical analyses using one-way analysis of variance (ANOVA) followed by Bonferroni’s Multiple Comparison Tests.


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







