
High-Throughput Screening Campaign Identified a Potential Small Molecule RXFP3/4 Agonist

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
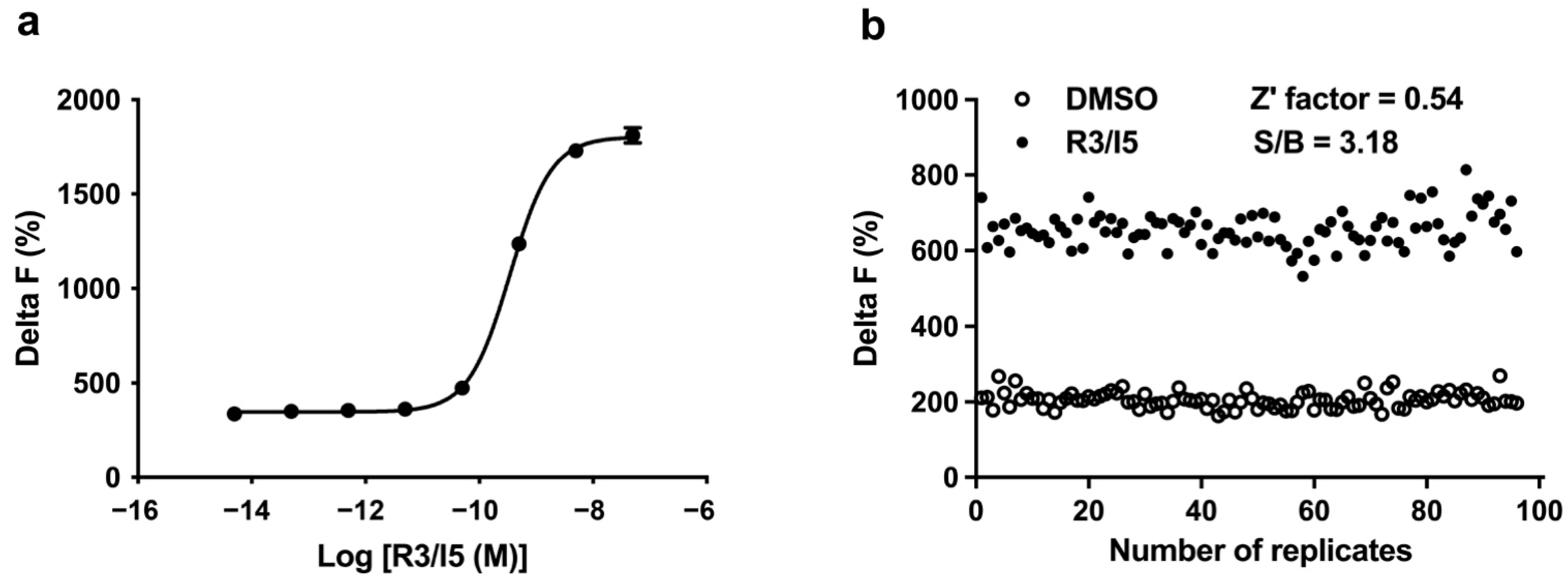
2.1. Assay Validation
2.2. Assay Performance
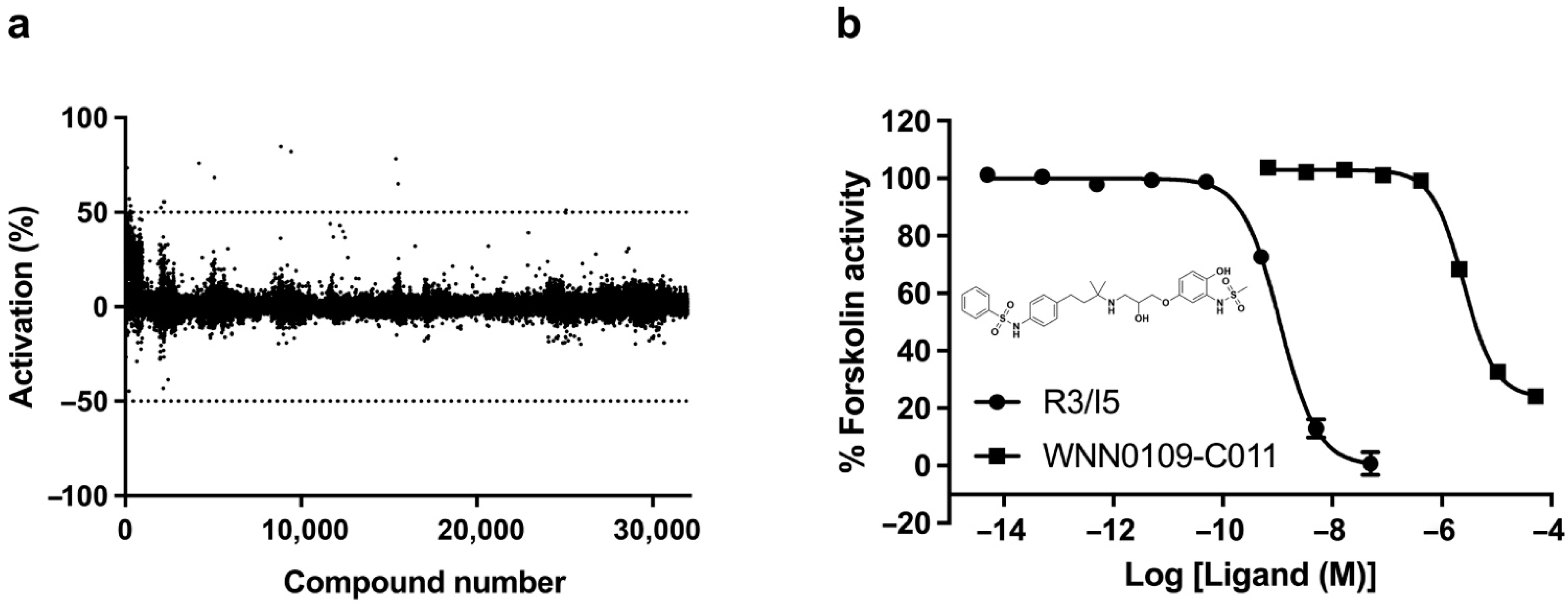
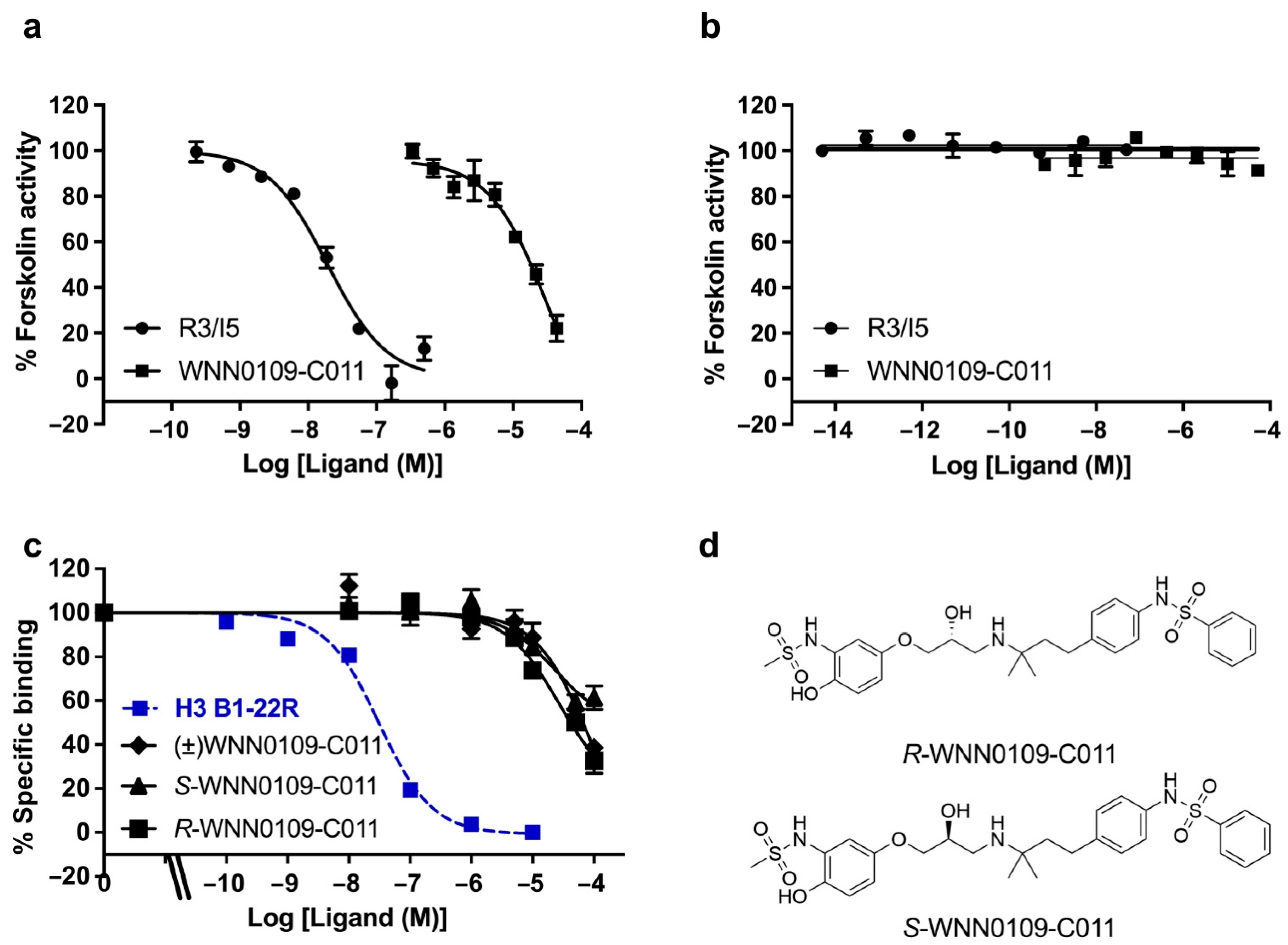
2.3. Lead Identification
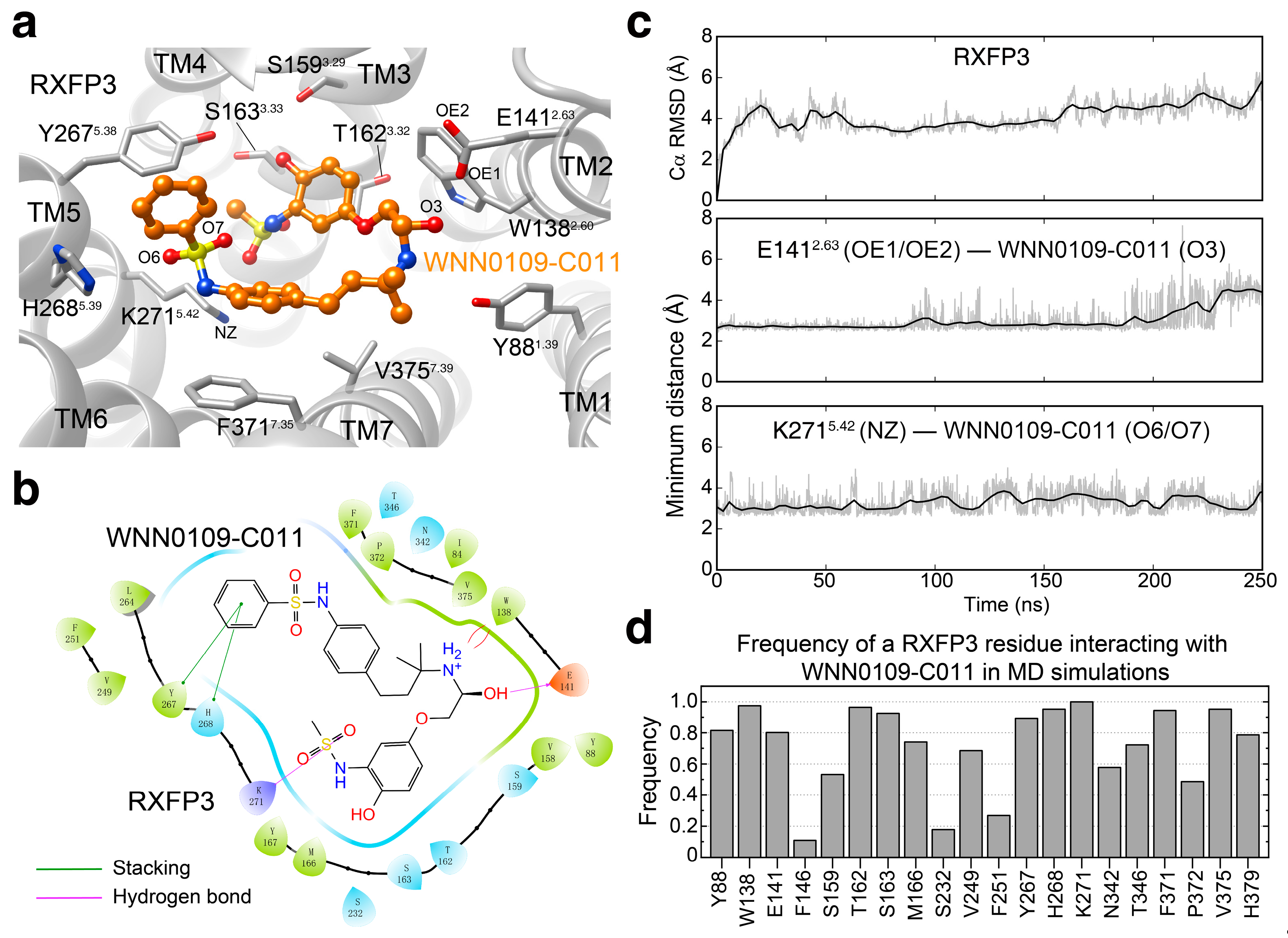
2.4. Binding Pose Prediction
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Compound Library
4.4. HTS Campaign
4.5. cAMP Assay
4.6. Europium-Labelled H3 B1-22R Competition Binding
4.7. Molecular Docking
4.8. Molecular Dynamics Simulation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsumoto, M.; Kamohara, M.; Sugimoto, T.; Hidaka, K.; Takasaki, J.; Saito, T.; Okada, M.; Yamaguchi, T.; Furuichi, K. The novel G-protein coupled receptor SALPR shares sequence similarity with somatostatin and angiotensin receptors. Gene 2000, 248, 183–189. [Google Scholar] [CrossRef]
- Fredriksson, R.; Lagerstrom, M.C.; Lundin, L.G.; Schioth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathgate, R.A.; Ivell, R.; Sanborn, B.M.; Sherwood, O.D.; Summers, R.J. International Union of Pharmacology LVII: Recommendations for the nomenclature of receptors for relaxin family peptides. Pharmacol. Rev. 2006, 58, 7–31. [Google Scholar] [CrossRef]
- Liu, C.; Eriste, E.; Sutton, S.; Chen, J.; Roland, B.; Kuei, C.; Farmer, N.; Jornvall, H.; Sillard, R.; Lovenberg, T.W. Identification of relaxin-3/INSL7 as an endogenous ligand for the orphan G-protein-coupled receptor GPCR135. J. Biol. Chem. 2003, 278, 50754–50764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathgate, R.A.; Halls, M.L.; van der Westhuizen, E.T.; Callander, G.E.; Kocan, M.; Summers, R.J. Relaxin family peptides and their receptors. Physiol. Rev. 2013, 93, 405–480. [Google Scholar] [CrossRef]
- Ma, S.; Bonaventure, P.; Ferraro, T.; Shen, P.J.; Burazin, T.C.; Bathgate, R.A.; Liu, C.; Tregear, G.W.; Sutton, S.W.; Gundlach, A.L. Relaxin-3 in GABA projection neurons of nucleus incertus suggests widespread influence on forebrain circuits via G-protein-coupled receptor-135 in the rat. Neuroscience 2007, 144, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Sutton, S.W.; Bonaventure, P.; Kuei, C.; Roland, B.; Chen, J.; Nepomuceno, D.; Lovenberg, T.W.; Liu, C. Distribution of G-protein-coupled receptor (GPCR)135 binding sites and receptor mRNA in the rat brain suggests a role for relaxin-3 in neuroendocrine and sensory processing. Neuroendocrinology 2004, 80, 298–307. [Google Scholar] [CrossRef]
- Smith, C.M.; Shen, P.J.; Banerjee, A.; Bonaventure, P.; Ma, S.; Bathgate, R.A.; Sutton, S.W.; Gundlach, A.L. Distribution of relaxin-3 and RXFP3 within arousal, stress, affective, and cognitive circuits of mouse brain. J. Comp. Neurol. 2010, 518, 4016–4045. [Google Scholar] [CrossRef]
- Ma, S.; Smith, C.M.; Blasiak, A.; Gundlach, A.L. Distribution, physiology and pharmacology of relaxin-3/RXFP3 systems in brain. Br. J. Pharmacol. 2017, 174, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Shen, P.J.; Sang, Q.; Lanciego, J.L.; Gundlach, A.L. Distribution of relaxin-3 mRNA and immunoreactivity and RXFP3-binding sites in the brain of the macaque, Macaca fascicularis. Ann. N. Y. Acad. Sci. 2009, 1160, 256–258. [Google Scholar] [CrossRef]
- Liu, C.; Lovenberg, T.W. Relaxin-3, INSL5, and their receptors. Results Probl. Cell Differ. 2008, 46, 213–237. [Google Scholar] [CrossRef] [PubMed]
- Bathgate, R.A.; Samuel, C.S.; Burazin, T.C.; Layfield, S.; Claasz, A.A.; Reytomas, I.G.; Dawson, N.F.; Zhao, C.; Bond, C.; Summers, R.J.; et al. Human relaxin gene 3 (H3) and the equivalent mouse relaxin (M3) gene. Novel members of the relaxin peptide family. J. Biol. Chem. 2002, 277, 1148–1157. [Google Scholar] [CrossRef] [Green Version]
- Burazin, T.C.; Bathgate, R.A.; Macris, M.; Layfield, S.; Gundlach, A.L.; Tregear, G.W. Restricted, but abundant, expression of the novel rat gene-3 (R3) relaxin in the dorsal tegmental region of brain. J. Neurochem. 2002, 82, 1553–1557. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Iijima, N.; Miyamoto, Y.; Fukusumi, S.; Itoh, Y.; Ozawa, H.; Ibata, Y. Neurons expressing relaxin 3/INSL 7 in the nucleus incertus respond to stress. Eur. J. Neurosci. 2005, 21, 1659–1670. [Google Scholar] [CrossRef]
- Ma, S.; Sang, Q.; Lanciego, J.L.; Gundlach, A.L. Localization of relaxin-3 in brain of Macaca fascicularis: Identification of a nucleus incertus in primate. J. Comp. Neurol. 2009, 517, 856–872. [Google Scholar] [CrossRef]
- van der Westhuizen, E.T.; Werry, T.D.; Sexton, P.M.; Summers, R.J. The relaxin family peptide receptor 3 activates extracellular signal-regulated kinase 1/2 through a protein kinase C-dependent mechanism. Mol. Pharmacol. 2007, 71, 1618–1629. [Google Scholar] [CrossRef]
- van der Westhuizen, E.T.; Christopoulos, A.; Sexton, P.M.; Wade, J.D.; Summers, R.J. H2 relaxin is a biased ligand relative to H3 relaxin at the relaxin family peptide receptor 3 (RXFP3). Mol. Pharmacol. 2010, 77, 759–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocan, M.; Sarwar, M.; Hossain, M.A.; Wade, J.D.; Summers, R.J. Signalling profiles of H3 relaxin, H2 relaxin and R3(BDelta23-27)R/I5 acting at the relaxin family peptide receptor 3 (RXFP3). Br. J. Pharmacol. 2014, 171, 2827–2841. [Google Scholar] [CrossRef] [Green Version]
- Hosken, I.T.; Sutton, S.W.; Smith, C.M.; Gundlach, A.L. Relaxin-3 receptor (Rxfp3) gene knockout mice display reduced running wheel activity: Implications for role of relaxin-3/RXFP3 signalling in sustained arousal. Behav. Brain Res. 2015, 278, 167–175. [Google Scholar] [CrossRef]
- Watanabe, Y.; Tsujimura, A.; Takao, K.; Nishi, K.; Ito, Y.; Yasuhara, Y.; Nakatomi, Y.; Yokoyama, C.; Fukui, K.; Miyakawa, T.; et al. Relaxin-3-deficient mice showed slight alteration in anxiety-related behavior. Front. Behav. Neurosci. 2011, 5, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.M.; Hosken, I.T.; Sutton, S.W.; Lawrence, A.J.; Gundlach, A.L. Relaxin-3 null mutation mice display a circadian hypoactivity phenotype. Genes Brain Behav. 2012, 11, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Blasiak, A.; Ganella, D.E.; Chua, B.E.; Layfield, S.; Bathgate, R.A.; Gundlach, A.L. Viral-mediated delivery of an RXFP3 agonist into brain promotes arousal in mice. Ital. J. Anat. Embryol. 2013, 118, 42–46. [Google Scholar]
- McGowan, B.M.; Stanley, S.A.; Smith, K.L.; White, N.E.; Connolly, M.M.; Thompson, E.L.; Gardiner, J.V.; Murphy, K.G.; Ghatei, M.A.; Bloom, S.R. Central relaxin-3 administration causes hyperphagia in male Wistar rats. Endocrinology 2005, 146, 3295–3300. [Google Scholar] [CrossRef] [PubMed]
- McGowan, B.M.; Stanley, S.A.; Smith, K.L.; Minnion, J.S.; Donovan, J.; Thompson, E.L.; Patterson, M.; Connolly, M.M.; Abbott, C.R.; Small, C.J.; et al. Effects of acute and chronic relaxin-3 on food intake and energy expenditure in rats. Regul. Pept. 2006, 136, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Hida, T.; Takahashi, E.; Shikata, K.; Hirohashi, T.; Sawai, T.; Seiki, T.; Tanaka, H.; Kawai, T.; Ito, O.; Arai, T.; et al. Chronic intracerebroventricular administration of relaxin-3 increases body weight in rats. J. Recept. Signal. Transduct. Res. 2006, 26, 147–158. [Google Scholar] [CrossRef]
- Ganella, D.E.; Callander, G.E.; Ma, S.; Bye, C.R.; Gundlach, A.L.; Bathgate, R.A. Modulation of feeding by chronic rAAV expression of a relaxin-3 peptide agonist in rat hypothalamus. Gene Ther. 2013, 20, 703–716. [Google Scholar] [CrossRef] [PubMed]
- de Avila, C.; Chometton, S.; Lenglos, C.; Calvez, J.; Gundlach, A.L.; Timofeeva, E. Differential effects of relaxin-3 and a selective relaxin-3 receptor agonist on food and water intake and hypothalamic neuronal activity in rats. Behav. Brain Res. 2018, 336, 135–144. [Google Scholar] [CrossRef] [Green Version]
- DeChristopher, B.; Park, S.H.; Vong, L.; Bamford, D.; Cho, H.H.; Duvadie, R.; Fedolak, A.; Hogan, C.; Honda, T.; Pandey, P.; et al. Discovery of a small molecule RXFP3/4 agonist that increases food intake in rats upon acute central administration. Bioorg. Med. Chem. Lett. 2019, 29, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Shabanpoor, F.; Akhter Hossain, M.; Ryan, P.J.; Belgi, A.; Layfield, S.; Kocan, M.; Zhang, S.; Samuel, C.S.; Gundlach, A.L.; Bathgate, R.A.; et al. Minimization of human relaxin-3 leading to high-affinity analogues with increased selectivity for relaxin-family peptide 3 receptor (RXFP3) over RXFP1. J. Med. Chem. 2012, 55, 1671–1681. [Google Scholar] [CrossRef]
- Kuei, C.; Sutton, S.; Bonaventure, P.; Pudiak, C.; Shelton, J.; Zhu, J.; Nepomuceno, D.; Wu, J.; Chen, J.; Kamme, F.; et al. R3(BDelta23 27)R/I5 chimeric peptide, a selective antagonist for GPCR135 and GPCR142 over relaxin receptor LGR7: In vitro and in vivo characterization. J. Biol. Chem. 2007, 282, 25425–25435. [Google Scholar] [CrossRef] [Green Version]
- Haugaard-Kedstrom, L.M.; Shabanpoor, F.; Hossain, M.A.; Clark, R.J.; Ryan, P.J.; Craik, D.J.; Gundlach, A.L.; Wade, J.D.; Bathgate, R.A.; Rosengren, K.J. Design, synthesis, and characterization of a single-chain peptide antagonist for the relaxin-3 receptor RXFP3. J. Am. Chem. Soc. 2011, 133, 4965–4974. [Google Scholar] [CrossRef] [PubMed]
- Albert-Gasco, H.; Garcia-Aviles, A.; Moustafa, S.; Sanchez-Sarasua, S.; Gundlach, A.L.; Olucha-Bordonau, F.E.; Sanchez-Perez, A.M. Central relaxin-3 receptor (RXFP3) activation increases ERK phosphorylation in septal cholinergic neurons and impairs spatial working memory. Brain Struct. Funct. 2017, 222, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, J.R.; Rajkumar, R.; Jayakody, T.; Marwari, S.; Hong, J.M.; Ma, S.; Gundlach, A.L.; Lai, M.K.P.; Dawe, G.S. Relaxin’ the brain: A case for targeting the nucleus incertus network and relaxin-3/RXFP3 system in neuropsychiatric disorders. Br. J. Pharmacol. 2017, 174, 1061–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.M.; Walker, A.W.; Hosken, I.T.; Chua, B.E.; Zhang, C.; Haidar, M.; Gundlach, A.L. Relaxin-3/RXFP3 networks: An emerging target for the treatment of depression and other neuropsychiatric diseases? Front. Pharmacol. 2014, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Ryan, P.J.; Buchler, E.; Shabanpoor, F.; Hossain, M.A.; Wade, J.D.; Lawrence, A.J.; Gundlach, A.L. Central relaxin-3 receptor (RXFP3) activation decreases anxiety- and depressive-like behaviours in the rat. Behav. Brain Res. 2013, 244, 142–151. [Google Scholar] [CrossRef]
- Hossain, M.A.; Rosengren, K.J.; Haugaard-Jonsson, L.M.; Zhang, S.; Layfield, S.; Ferraro, T.; Daly, N.L.; Tregear, G.W.; Wade, J.D.; Bathgate, R.A. The A-chain of human relaxin family peptides has distinct roles in the binding and activation of the different relaxin family peptide receptors. J. Biol. Chem. 2008, 283, 17287–17297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Chen, J.; Kuei, C.; Sutton, S.; Nepomuceno, D.; Bonaventure, P.; Lovenberg, T.W. Relaxin-3/insulin-like peptide 5 chimeric peptide, a selective ligand for G protein-coupled receptor (GPCR)135 and GPCR142 over leucine-rich repeat-containing G protein-coupled receptor 7. Mol. Pharmacol. 2005, 67, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Bathgate, R.A.; Rosengren, K.J.; Shabanpoor, F.; Zhang, S.; Lin, F.; Tregear, G.W.; Wade, J.D. The structural and functional role of the B-chain C-terminal arginine in the relaxin-3 peptide antagonist, R3(BDelta23-27)R/I5. Chem. Biol. Drug Des. 2009, 73, 46–52. [Google Scholar] [CrossRef]
- Wei, D.; Hu, M.J.; Shao, X.X.; Wang, J.H.; Nie, W.H.; Liu, Y.L.; Xu, Z.G.; Guo, Z.Y. Development of a selective agonist for relaxin family peptide receptor 3. Sci. Rep. 2017, 7, 3230. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Jaimes, L.; Sutton, S.W.; Nepomuceno, D.; Motley, S.T.; Cik, M.; Stocking, E.; Shoblock, J.; Bonaventure, P. In vitro pharmacological characterization of RXFP3 allosterism: An example of probe dependency. PLoS ONE 2012, 7, e30792. [Google Scholar] [CrossRef]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A Simple Statistical Parameter for Use in Evaluation and Validation of High Throughput Screening Assays. J. Biomol. Screen 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.-Y.; Lin, L.; Cai, X.-Q.; Dai, A.-T.; Zhu, Y.; Li, J.; Liu, Q.; Yang, D.-H.; Bathgate, R.A.D.; Wang, M.-W. High-throughput screening campaign identifies a small molecule agonist of the relaxin family peptide receptor 4. Acta Pharmacol. Sin. 2020, 41, 1328–1336. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Weinstein, H. [19] Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. In Methods in Neurosciences; Receptor Molecular Biology; Academic Press: San Diego, CA, USA, 1995; pp. 366–428. [Google Scholar]
- Van der Westhuizen, E.T.; Sexton, P.M.; Bathgate, R.A.; Summers, R.J. Responses of GPCR135 to human gene 3 (H3) relaxin in CHO-K1 cells determined by microphysiometry. Ann. N. Y. Acad. Sci. 2005, 1041, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Belgi, A.; Hossain, M.A.; Shabanpoor, F.; Chan, L.; Zhang, S.; Bathgate, R.A.; Tregear, G.W.; Wade, J.D. Structure and function relationship of murine insulin-like peptide 5 (INSL5): Free C-terminus is essential for RXFP4 receptor binding and activation. Biochemistry 2011, 50, 8352–8361. [Google Scholar] [CrossRef]
- Chinese National Compound Library. Available online: https://www.cncl.org.cn (accessed on 1 January 2020).
- Haugaard-Kedstrom, L.M.; Wong, L.L.; Bathgate, R.A.; Rosengren, K.J. Synthesis and pharmacological characterization of a europium-labelled single-chain antagonist for binding studies of the relaxin-3 receptor RXFP3. Amino Acids 2015, 47, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- GPCRdb. Available online: https://gpcrdb.org/ (accessed on 20 November 2021).
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Davila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M.; et al. CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [Green Version]
- Guvench, O.; Mallajosyula, S.S.; Raman, E.P.; Hatcher, E.; Vanommeslaeghe, K.; Foster, T.J.; Jamison, F.W., 2nd; Mackerell, A.D., Jr. CHARMM additive all-atom force field for carbohydrate derivatives and its utility in polysaccharide and carbohydrate-protein modeling. J. Chem. Theory Comput. 2011, 7, 3162–3180. [Google Scholar] [CrossRef] [Green Version]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Aoki, K.M.; Yonezawa, F. Constant-pressure molecular-dynamics simulations of the crystal-smectic transition in systems of soft parallel spherocylinders. Phys. Rev. A 1992, 46, 6541–6549. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, G.; Feng, Y.; Cai, X.; Zhou, C.; Shao, L.; Chen, Y.; Chen, L.; Liu, Q.; Zhou, Q.; Bathgate, R.A.D.; et al. High-Throughput Screening Campaign Identified a Potential Small Molecule RXFP3/4 Agonist. Molecules 2021, 26, 7511. https://doi.org/10.3390/molecules26247511
Lin G, Feng Y, Cai X, Zhou C, Shao L, Chen Y, Chen L, Liu Q, Zhou Q, Bathgate RAD, et al. High-Throughput Screening Campaign Identified a Potential Small Molecule RXFP3/4 Agonist. Molecules. 2021; 26(24):7511. https://doi.org/10.3390/molecules26247511
Chicago/Turabian StyleLin, Guangyao, Yang Feng, Xiaoqing Cai, Caihong Zhou, Lijun Shao, Yan Chen, Linhai Chen, Qing Liu, Qingtong Zhou, Ross A.D. Bathgate, and et al. 2021. "High-Throughput Screening Campaign Identified a Potential Small Molecule RXFP3/4 Agonist" Molecules 26, no. 24: 7511. https://doi.org/10.3390/molecules26247511