
Attenuation of Tumor Development in Mammary Carcinoma Rats by Theacrine, an Antagonist of Adenosine 2A Receptor


Abstract
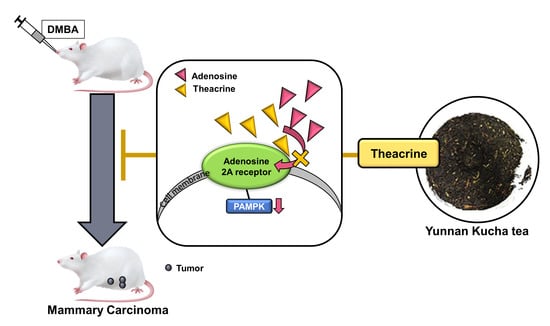
:
1. Introduction
2. Results
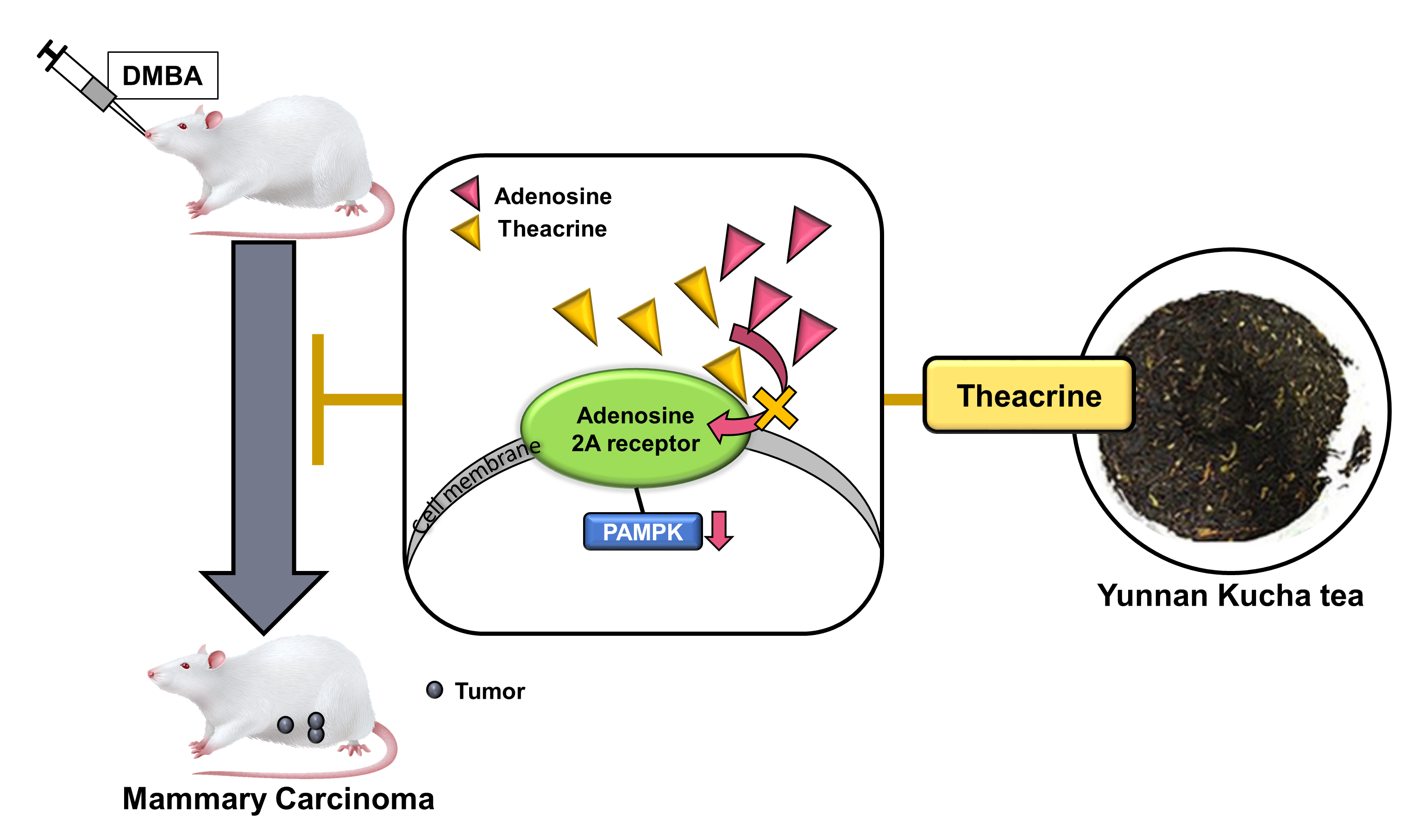
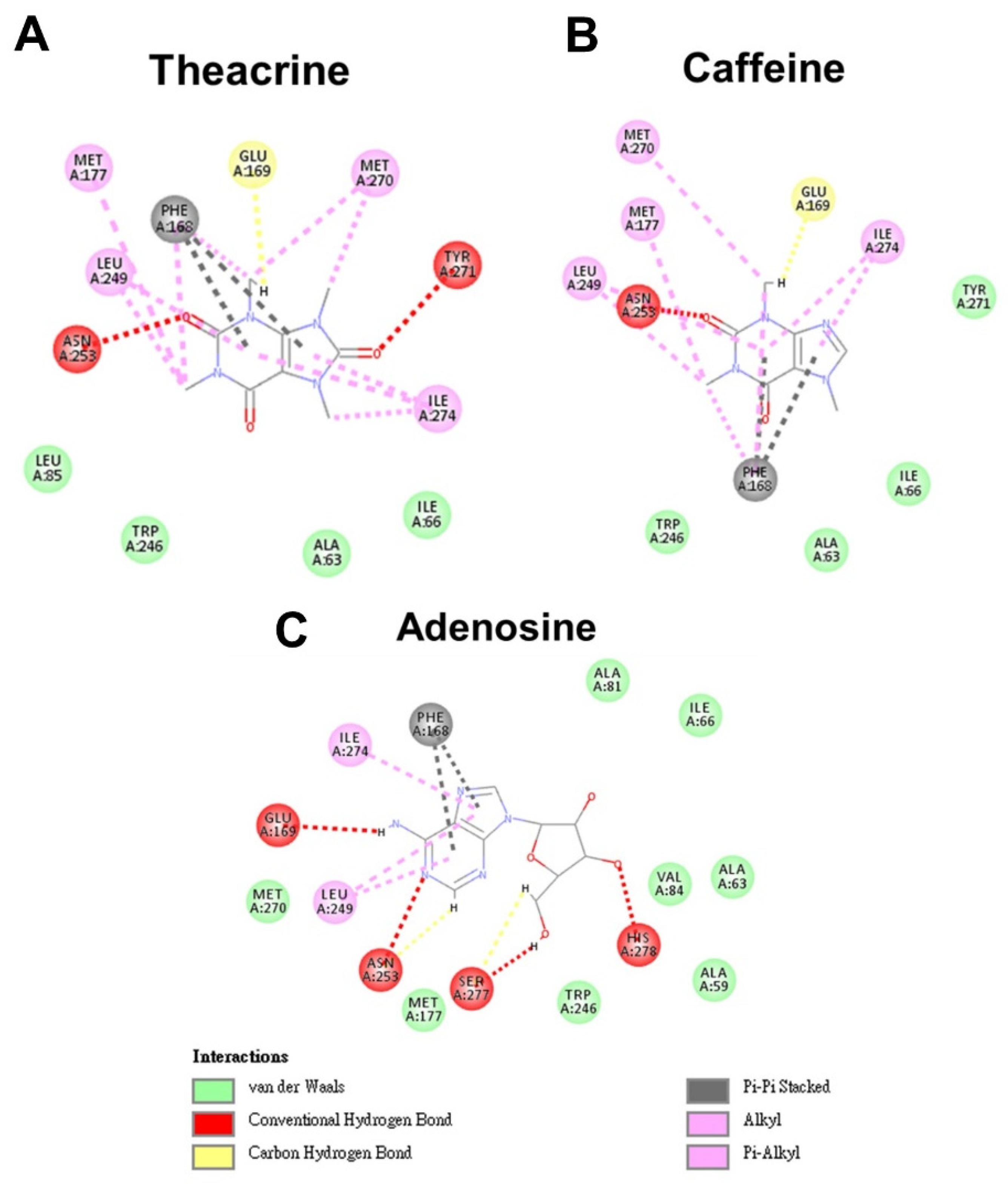
2.1. Molecular Modeling of Theacrine, Caffeine and Adenosine Docking to the Adenosine 2A Receptor
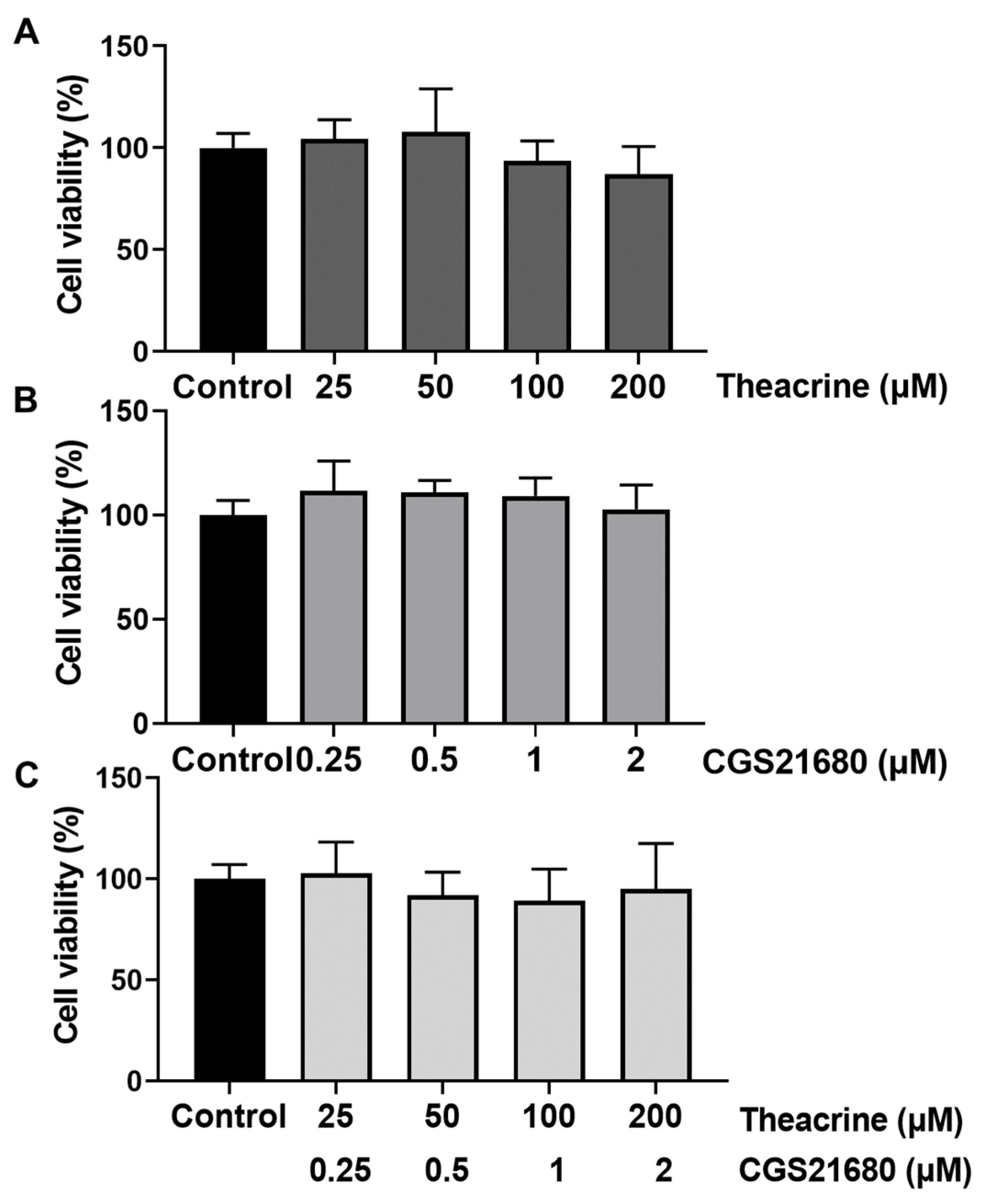
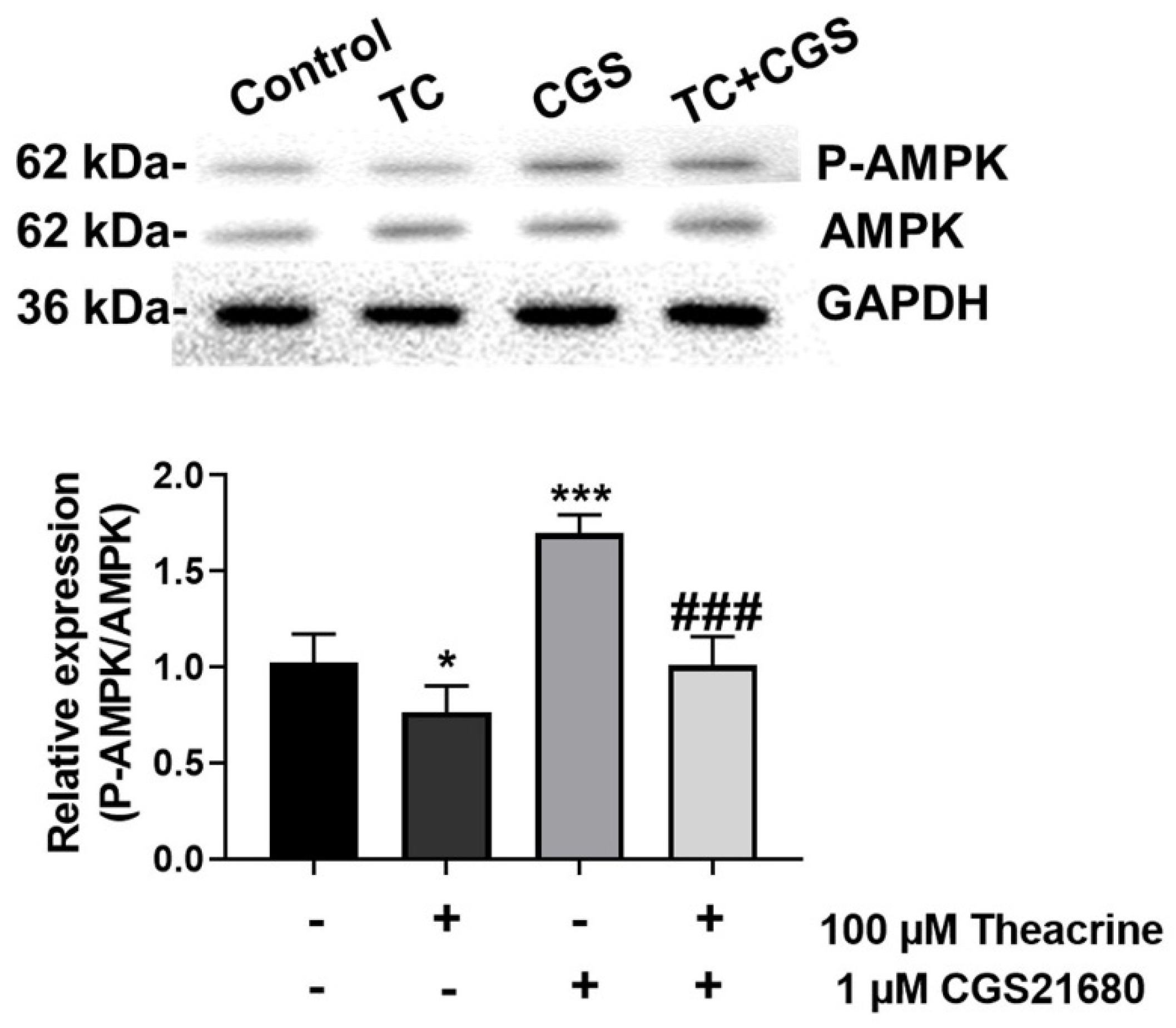
2.2. Detecting Theacrine as an Antagonist of the Adenosine 2A Receptor
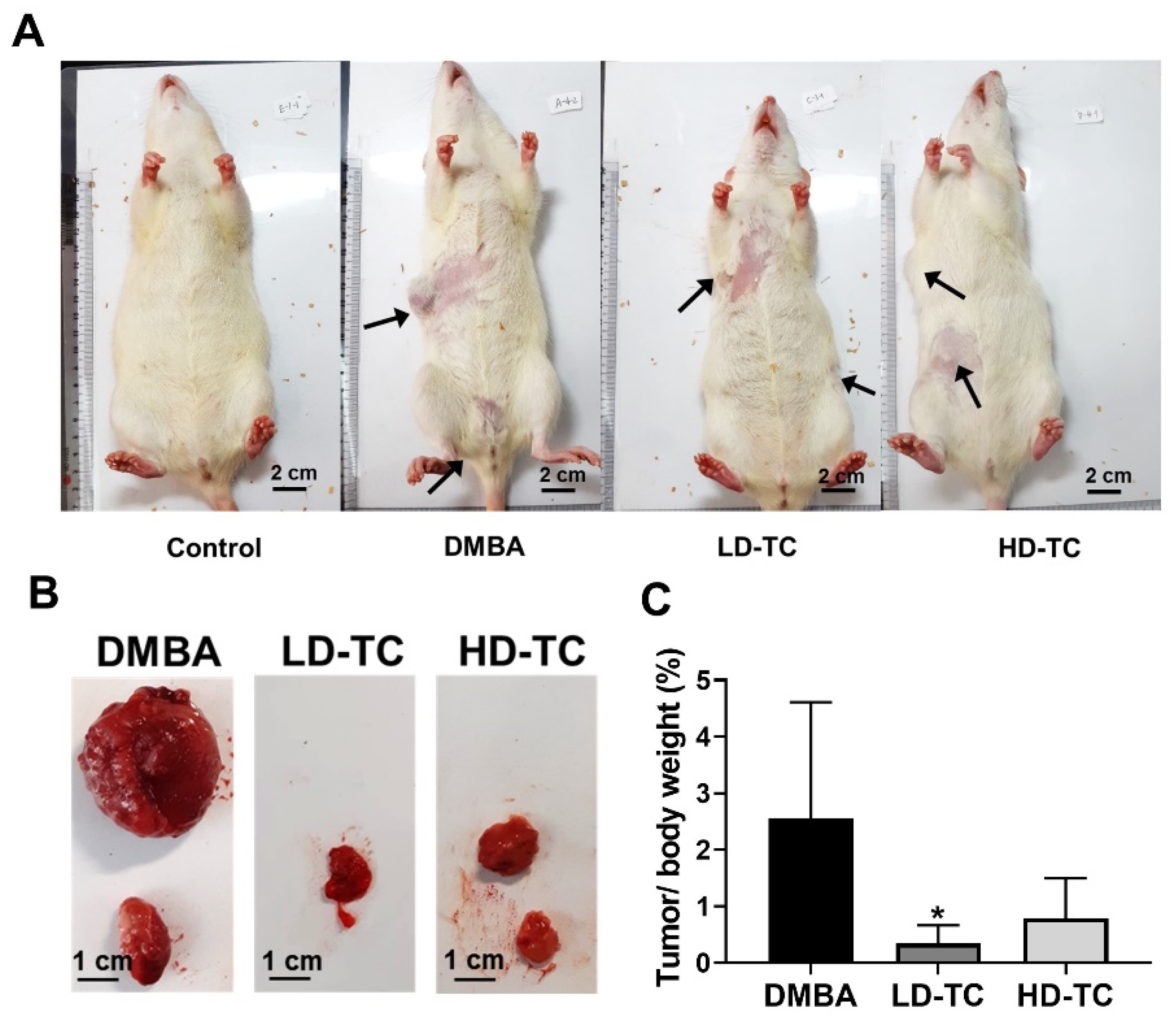
2.3. Effects of Theacrine on DMBA-Induced Mammary Carcinoma in Rats
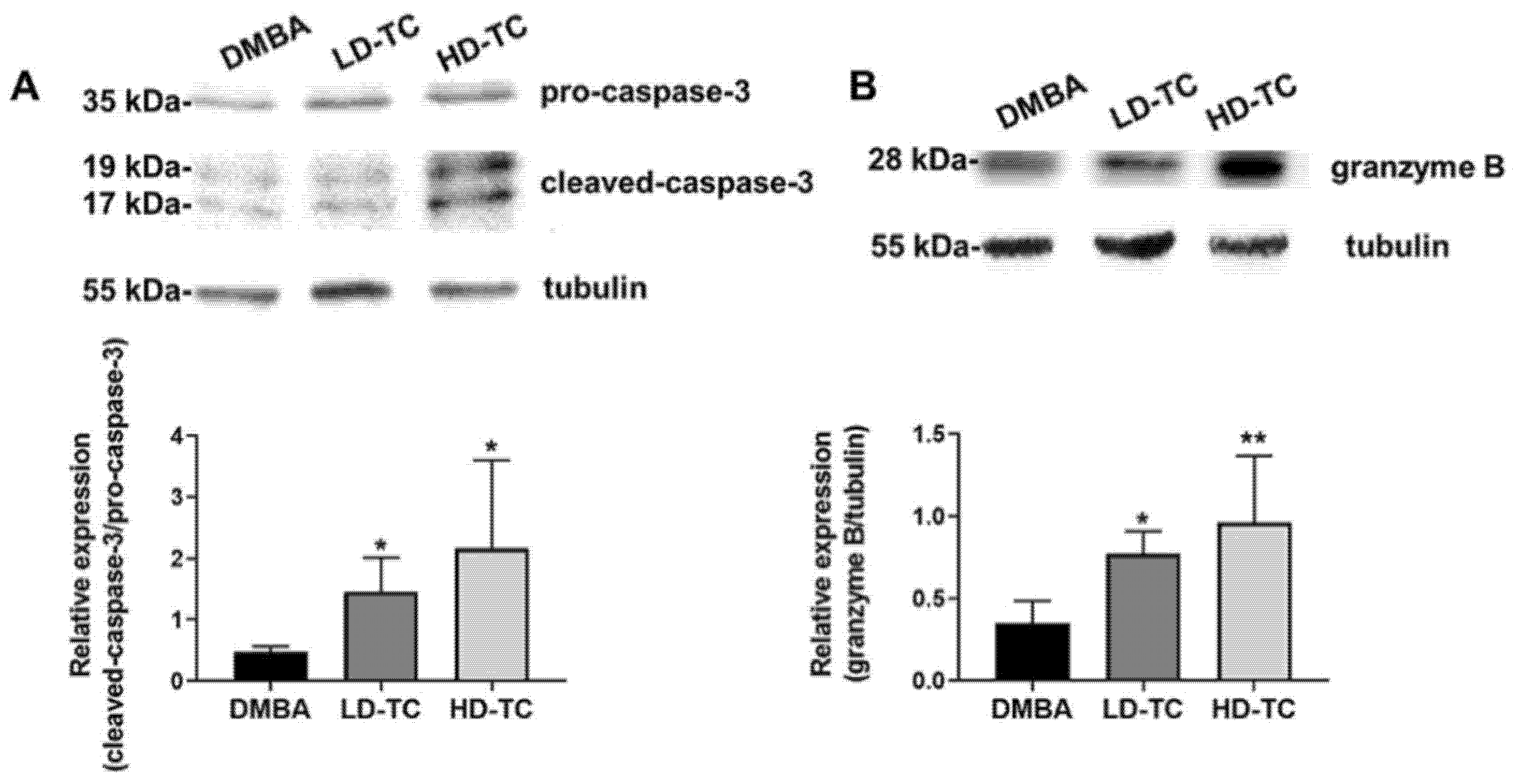
2.4. Elevation of Programmed Cell Death in DMBA-Induced Tumor by Theacrine
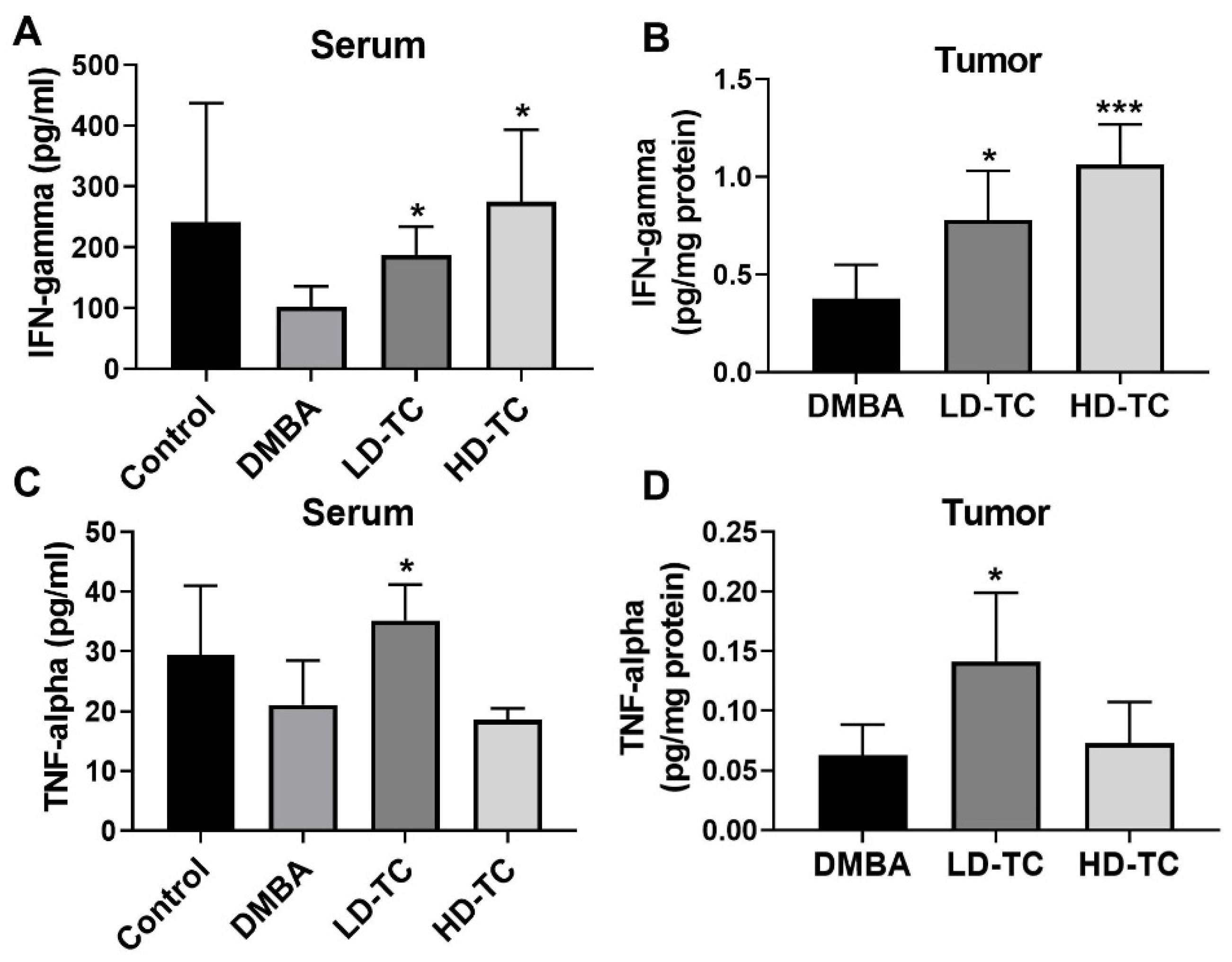
2.5. Effects of Theacrine on the Levels of IFN-γ and TNF-α in DMBA-Treated Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Homology Modeling and Docking
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Animals
4.6. Estimation of Body and Tumor Parameters
4.7. Enzyme Linked Immune Sorbent Assay for Proinflammatory Cytokines
4.8. Western Blotting
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Joshi, H.; Press, M.F. 22—Molecular Oncology of Breast Cancer. In The Breast, 5th ed.; Bland, K.I., Copeland, E.M., Klimberg, V.S., Gradishar, W.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 282–307.e5. [Google Scholar]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Korde, L.A.; Somerfield, M.R.; Carey, L.A.; Crews, J.R.; Denduluri, N.; Hwang, E.S.; Khan, S.A.; Loibl, S.; Morris, E.A.; Perez, A.; et al. Neoadjuvant Chemotherapy, Endocrine Therapy, and Targeted Therapy for Breast Cancer: ASCO Guideline. J. Clin. Oncol. 2021, 39, 1485–1505. [Google Scholar] [CrossRef] [PubMed]
- García-Aranda, M.; Redondo, M. Immunotherapy: A Challenge of Breast Cancer Treatment. Cancers 2019, 11, 1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.; Gatti-Mays, M.E.; Kalinsky, K.; Korde, L.A.; Sharon, E.; Amiri-Kordestani, L.; Bear, H.; McArthur, H.L.; Frank, E.; Perlmutter, J.; et al. Current Landscape of Immunotherapy in Breast Cancer: A Review. JAMA Oncol. 2019, 5, 1205–1214. [Google Scholar] [CrossRef]
- Marin-Acevedo, J.A.; Dholaria, B.; Soyano, A.E.; Knutson, K.L.; Chumsri, S.; Lou, Y. Next generation of immune checkpoint therapy in cancer: New developments and challenges. J. Hematol. Oncol. 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Arab, S.; Hadjati, J. Adenosine Blockage in Tumor Microenvironment and Improvement of Cancer Immunotherapy. Immune Netw. 2019, 19, e23. [Google Scholar] [CrossRef] [PubMed]
- Allard, B.; Beavis, P.A.; Darcy, P.K.; Stagg, J. Immunosuppressive activities of adenosine in cancer. Curr. Opin. Pharmacol. 2016, 29, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Bencivenni, S.; Battistello, E.; Vincenzi, F.; Colotta, V.; Catarzi, D.; Varano, F.; Merighi, S.; Borea, P.A.; Varani, K. Inhibition of A(2A) Adenosine Receptor Signaling in Cancer Cells Proliferation by the Novel Antagonist TP455. Front. Pharmacol. 2017, 8, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borodovsky, A.; Barbon, C.M.; Wang, Y.; Ye, M.; Prickett, L.; Chandra, D.; Shaw, J.; Deng, N.; Sachsenmeier, K.; Clarke, J.D.; et al. Small molecule AZD4635 inhibitor of A(2A)R signaling rescues immune cell function including CD103(+) dendritic cells enhancing anti-tumor immunity. J. Immunother. Cancer 2020, 8, 603. [Google Scholar] [CrossRef]
- Eini, H.; Frishman, V.; Yulzari, R.; Kachko, L.; Lewis, E.C.; Chaimovitz, C.; Douvdevani, A. Caffeine promotes anti-tumor immune response during tumor initiation: Involvement of the adenosine A2A receptor. Biochem. Pharmacol. 2015, 98, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.L.; Hsu, Y.J.; Chen, Y.; Wang, Y.W.; Huang, S.M. Theophylline exhibits anti-cancer activity via suppressing SRSF3 in cervical and breast cancer cell lines. Oncotarget 2017, 8, 101461–101474. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.R.; Kuo, P.C.; Li, Y.C.; Jhuo, C.F.; Hsu, W.L.; Tzen, J.T.C. Theacrine and strictinin, two major ingredients for the anti-influenza activity of Yunnan Kucha tea. J. Ethnopharmacol. 2020, 262, 113190. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Ye, X.; Bai, X.; He, J.; Li, T.; Zhang, J.; Zhang, W.; Xu, J. Theacrine: A purine alkaloid from Camellia assamica var. kucha with a hypnotic property via the adenosine system. Neurosci. Lett. 2017, 659, 48–53. [Google Scholar] [CrossRef]
- Sheng, Y.Y.; Xiang, J.; Wang, Z.S.; Jin, J.; Wang, Y.Q.; Li, Q.S.; Li, D.; Fang, Z.T.; Lu, J.L.; Ye, J.H.; et al. Theacrine From Camellia kucha and Its Health Beneficial Effects. Front. Nutr. 2020, 7, 596823. [Google Scholar] [CrossRef]
- Taylor, L.; Mumford, P.; Roberts, M.; Hayward, S.; Mullins, J.; Urbina, S.; Wilborn, C. Safety of TeaCrine®, a non-habituating, naturally-occurring purine alkaloid over eight weeks of continuous use. J. Int Soc. Sports Nutr. 2016, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.K.; Kurihara, H.; Zhao, L.; Yao, X.S. Theacrine, a special purine alkaloid with sedative and hypnotic properties from Cammelia assamica var. kucha in mice. J. Asian Nat. Prod. Res. 2007, 9, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.H.; Zhai, Y.J.; Wu, Y.P.; Xie, G.; Wang, G.E.; Mao, Z.F.; Hu, H.H.; Luo, X.H.; Sun, W.Y.; Liang, L.; et al. Theacrine, a Potent Antidepressant Purine Alkaloid from a Special Chinese Tea, Promotes Adult Hippocampal Neurogenesis in Stressed Mice. J. Agric. Food Chem. 2021, 69, 7016–7027. [Google Scholar] [CrossRef]
- Ko, J.H.; Yang, M.H.; Baek, S.H.; Nam, D.; Jung, S.H.; Ahn, K.S. Theacrine attenuates epithelial mesenchymal transition in human breast cancer MDA-MB-231 cells. Phytother. Res. 2019, 33, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Feduccia, A.A.; Wang, Y.; Simms, J.A.; Yi, H.Y.; Li, R.; Bjeldanes, L.; Ye, C.; Bartlett, S.E. Locomotor activation by theacrine, a purine alkaloid structurally similar to caffeine: Involvement of adenosine and dopamine receptors. Pharmacol. Biochem. Behav. 2012, 102, 241–248. [Google Scholar] [CrossRef]
- Sánchez-Melgar, A.; Albasanz, J.L.; Pallàs, M.; Martín, M. Adenosine Metabolism in the Cerebral Cortex from Several Mice Models during Aging. Int. J. Mol. Sci 2020, 21, 7300. [Google Scholar] [CrossRef]
- Veierød, M.B.; Thelle, D.S.; Laake, P. Diet and risk of cutaneous malignant melanoma: A prospective study of 50,757 Norwegian men and women. Int. J. Cancer 1997, 71, 600–604. [Google Scholar] [CrossRef]
- Tallis, J.; Clarke, N.; Morris, R.; Richardson, D.; Ellis, M.; Eyre, E.; Duncan, M.; Noon, M. The prevalence and practices of caffeine use as an ergogenic aid in English professional soccer. Biol. Sport 2021, 38, 525–534. [Google Scholar] [CrossRef]
- Zheng, X.Q.; Ye, C.X.; Kato, M.; Crozier, A.; Ashihara, H. Theacrine (1,3,7,9-tetramethyluric acid) synthesis in leaves of a Chinese tea, kucha (Camellia assamica var. kucha). Phytochemistry 2002, 60, 129–134. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Li, Y.F.; Wang, Y.; Tan, L.; Cao, Z.Q.; Xie, C.; Xie, G.; Gong, H.B.; Sun, W.Y.; Ouyang, S.H.; et al. Identification and characterization of N9-methyltransferase involved in converting caffeine into non-stimulatory theacrine in tea. Nat. Commun. 2020, 11, 1473. [Google Scholar] [CrossRef]
- Cekic, C.; Linden, J. Adenosine A2A receptors intrinsically regulate CD8+ T cells in the tumor microenvironment. Cancer Res. 2014, 74, 7239–7249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaakola, V.P.; Griffith, M.T.; Hanson, M.A.; Cherezov, V.; Chien, E.Y.; Lane, J.R.; Ijzerman, A.P.; Stevens, R.C. The 2.6 angstrom crystal structure of a human A2A adenosine receptor bound to an antagonist. Science 2008, 322, 1211–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, C.C.; Kong, L.R.; Chen, X.H.; Ma, Y.; Pan, X.X.; Zhang, Z.B.; Gao, P.J. A(2A) Receptor Activation Attenuates Hypertensive Cardiac Remodeling via Promoting Brown Adipose Tissue-Derived FGF21. Cell Metab. 2018, 28, 476–489. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Binding Energy (kJ/mol) | Absolute Energy (kJ/mol) | Complex Energy (kJ/mol) |
|---|---|---|---|
| Theacrine | −105.892 | 32.5403 | −18,472.1 |
| Caffeine | −101.077 | 27.6803 | −18,479.5 |
| Adenosine | −106.911 | 61.2889 | −18,555.9 |
| Parameter | Control x | DMBA | LD-TC | HD-TC |
|---|---|---|---|---|
| Tumor incidence (%) y | 0.0 | 66.7 | 66.7 | 66.7 |
| Body weight (g) | 330.52 ± 22.86 | 314.51 ± 24.74 | 315.48 ± 25.19 | 329.94 ± 20.26 |
| Tumor Volume | ||||
|---|---|---|---|---|
| Group | Day 84 | Day 91 | Day 98 | Day 102 |
| Control x | 0.00 y | 0.00 | 0.00 | 0.00 |
| DMBA | 1.38 ± 0.76 | 2.76 ± 1.53 | 4.13 ± 2.76 | 11.62 ± 7.60 |
| LD-TC | 0.57 ± 0.55 | 0.71 ± 0.75 *z | 0.96 ± 1.13 * | 3.46 ± 4.77 * |
| HD-TC | 0.87 ± 0.87 | 1.72 ± 1.86 | 2.37 ± 2.39 | 4.71 ± 3.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jhuo, C.-F.; Hsu, Y.-Y.; Chen, W.-Y.; Tzen, J.T.C. Attenuation of Tumor Development in Mammary Carcinoma Rats by Theacrine, an Antagonist of Adenosine 2A Receptor. Molecules 2021, 26, 7455. https://doi.org/10.3390/molecules26247455
Jhuo C-F, Hsu Y-Y, Chen W-Y, Tzen JTC. Attenuation of Tumor Development in Mammary Carcinoma Rats by Theacrine, an Antagonist of Adenosine 2A Receptor. Molecules. 2021; 26(24):7455. https://doi.org/10.3390/molecules26247455
Chicago/Turabian StyleJhuo, Cian-Fen, Yu-Yu Hsu, Wen-Ying Chen, and Jason T. C. Tzen. 2021. "Attenuation of Tumor Development in Mammary Carcinoma Rats by Theacrine, an Antagonist of Adenosine 2A Receptor" Molecules 26, no. 24: 7455. https://doi.org/10.3390/molecules26247455