
Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns

 , ,
, ,
Abstract
:1. Introduction
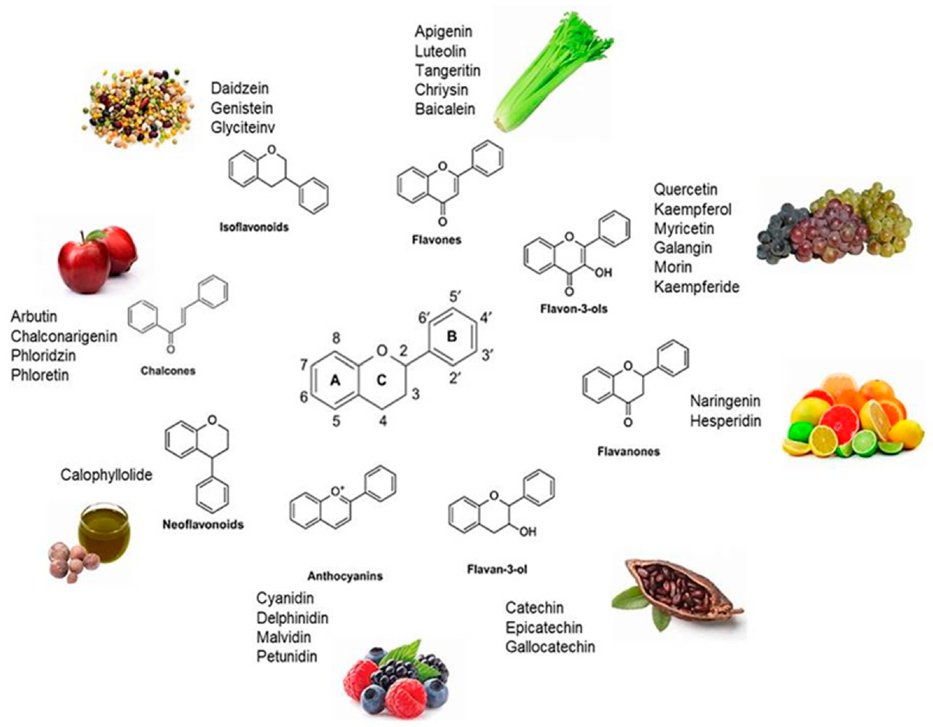
2. Dietary Flavonoids
2.1. Dietary Occurrence
2.2. Health Benefits, Medicinal Significance, and Nutraceutical Importance
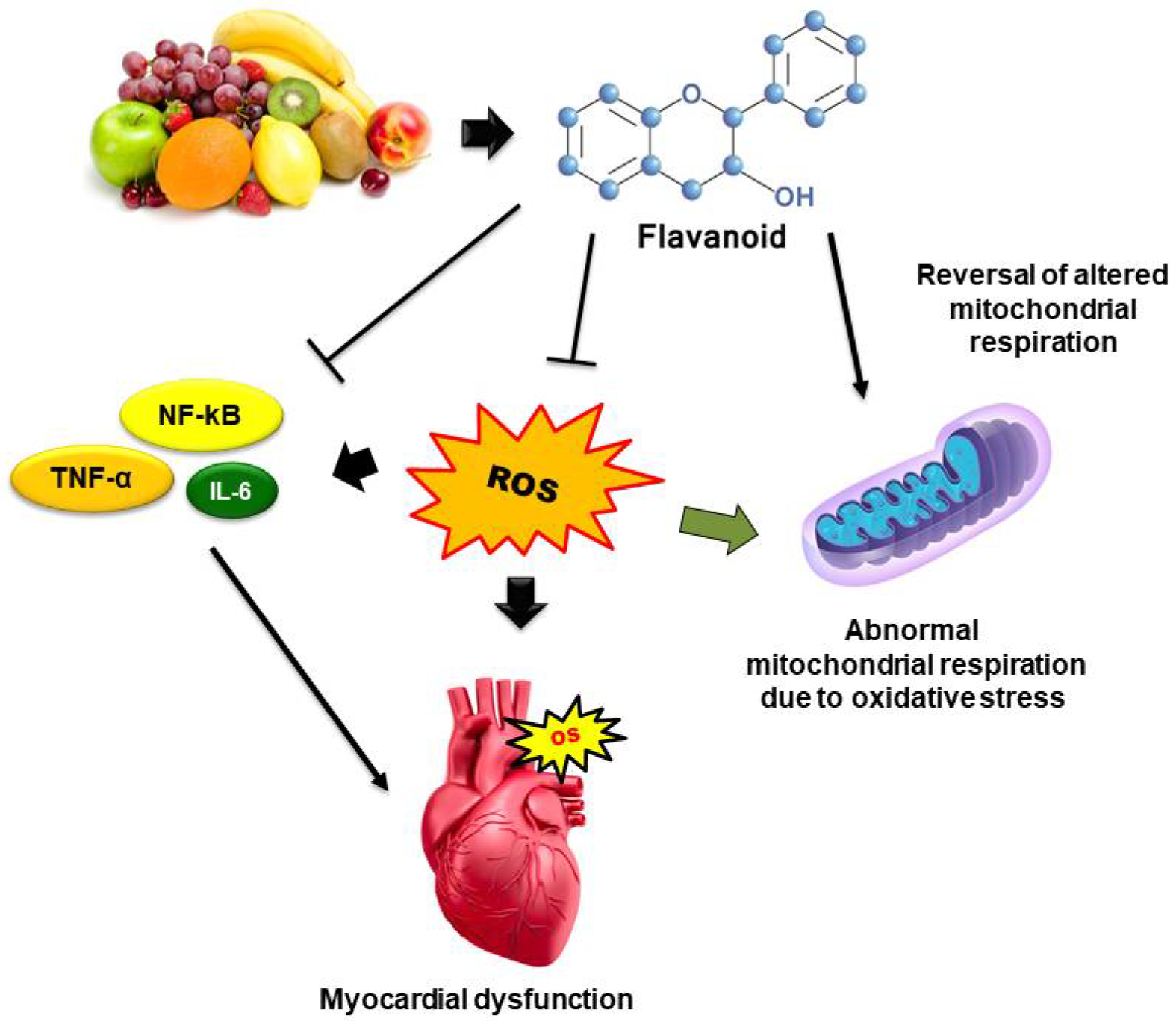
2.3. Antioxidant Potential of Dietary Flavonoids in OS-Induced CVDs
3. Cardioprotective Potential of Dietary Flavonoids
3.1. Dietary Flavonoids and Their Health Implications in CVDs
3.2. Cardioprotective Mechanisms of Dietary Flavonoids
3.2.1. ROS Scavenging Mechanism
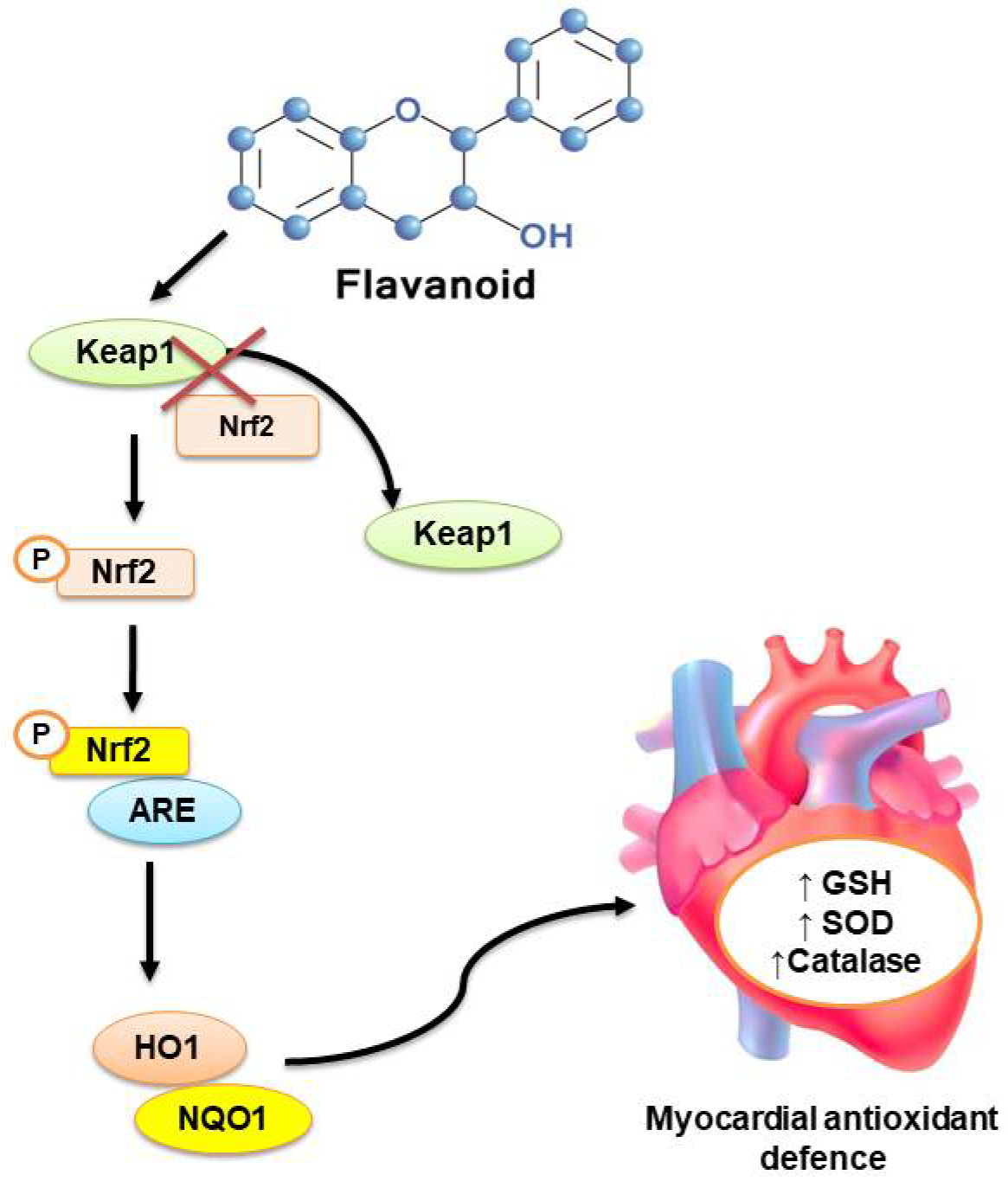
3.2.2. Intracellular Antioxidant Signaling Pathways
3.2.3. Counter-Inflammatory Pathways
3.2.4. Mitochondrial and Intracellular Pathways
4. Pharmacokinetic and Toxicological Issues
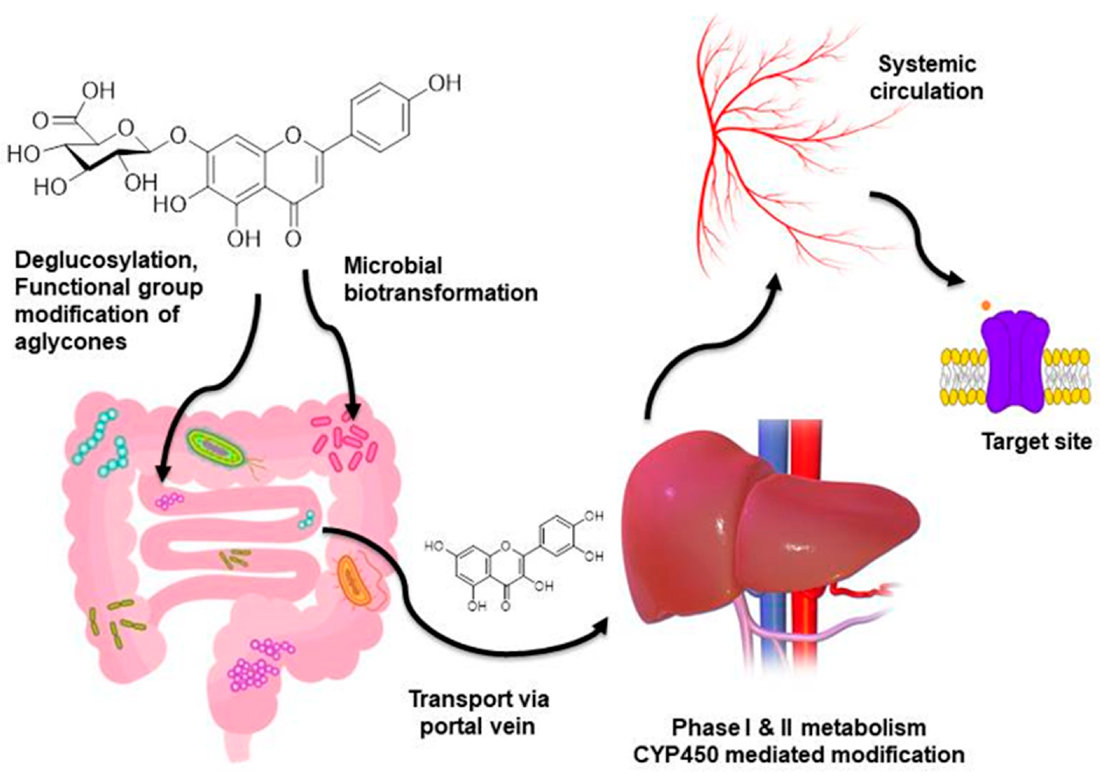
4.1. Bioavailability and Biotransformations of Dietary Flavonoids
4.2. Toxicities and Interactions with Drugs/Foods/Herbs
4.3. Strategies to Overcome Pharmacokinetic and Toxicological Limitations
5. Therapeutic Approaches and Future Drug Discovery
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Micha, R.; Peñalvo, J.L.; Cudhea, F.; Imamura, F.; Rehm, C.D.; Mozaffarian, D. Association between dietary factors and mortality from heart disease, stroke, and type 2 diabetes in the United States. J. Am. Med. Assoc. 2017, 317, 912–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, J.; Rodrigues, C.F.; Sharopov, F.; Docea, A.O.; Can Karaca, A.; Sharifi-Rad, M.; Kahveci Karıncaoglu, D.; Gülseren, G.; Şenol, E.; Demircan, E.; et al. Diet, lifestyle and cardiovascular diseases: Linking pathophysiology to cardioprotective effects of natural bioactive compounds. Int. J. Environ. Res. Public Health. 2020, 17, 2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Action Plan for the Prevention and Control of Noncommunicable Diseases 2013–2020; World Health Organization: Geneva, Switzerland, 2013; Available online: https://www.who.int/publications/i/item/9789241506236 (accessed on 16 March 2021).
- Yusuf, S.; Hawken, S.; Ôunpuu, S.; Dans, T.; Avezum, A.; Lanas, F.; McQueen, M.; Budaj, A.; Pais, P.; Varigos, J. Effect of potentially modifiable risk factors associated with myocardial infarction in 52 countries (the INTERHEART study): Case-control study. Lancet 2004, 364, 937–952. [Google Scholar] [CrossRef]
- Stewart, J.; Manmathan, G.; Wilkinson, P. Primary prevention of cardiovascular disease: A review of contemporary guidance and literature. JRSM Cardiovasc. Dis. 2017, 6, 2048004016687211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstädter, J.; Kröller-Schön, S.; Münzel, T. Vascular inflammation and oxidative stress: Major triggers for cardiovascular disease. Oxid. Med. Cell. Longev. 2019, 2019, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Kossmann, S.; Muenzel, T.; Daiber, A. Redox regulation of cardiovascular inflammation–Immunomodulatory function of mitochondrial and Nox-derived reactive oxygen and nitrogen species. Free Radic. Biol. Med. 2017, 109, 48–60. [Google Scholar] [CrossRef]
- Dikalova, A.E.; Bikineyeva, A.T.; Budzyn, K.; Nazarewicz, R.R.; McCann, L.; Lewis, W.; Harrison, D.G.; Dikalov, S.I. Therapeutic targeting of mitochondrial superoxide in hypertension. Circ. Res. 2010, 107, 106. [Google Scholar] [CrossRef] [Green Version]
- Gross, M. Flavonoids and cardiovascular disease. Pharm. Biol. 2004, 42, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Doughan, A.K.; Harrison, D.G.; Dikalov, S.I. Molecular mechanisms of angiotensin II–mediated mitochondrial dysfunction: Linking mitochondrial oxidative damage and vascular endothelial dysfunction. Circ. Res. 2008, 102, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Ábrigo, J.; Elorza, A.A.; Riedel, C.A.; Vilos, C.; Simon, F.; Cabrera, D.; Estrada, L.; Cabello-Verrugio, C. Role of oxidative stress as key regulator of muscle wasting during cachexia. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Yu, E.; Malik, V.S.; Hu, F.B. Cardiovascular disease prevention by diet modification: JACC health promotion series. J. Am. Coll. Cardiol. 2018, 72, 914–926. [Google Scholar] [CrossRef]
- Stranges, S.; Takeda, A.; Martin, N.; Rees, K. Cochrane corner: Does the Mediterranean-style diet help in the prevention of cardiovascular disease? Heart 2019, 105, 1691–1694. [Google Scholar] [CrossRef]
- Li, Y.; Hruby, A.; Bernstein, A.M.; Ley, S.H.; Wang, D.D.; Chiuve, S.E.; Sampson, L.; Rexrode, K.M.; Rimm, E.B.; Willett, W.C. Saturated fats compared with unsaturated fats and sources of carbohydrates in relation to risk of coronary heart disease: A prospective cohort study. J. Am. Coll. Cardiol. 2015, 66, 1538–1548. [Google Scholar] [CrossRef] [Green Version]
- He, F.J.; MacGregor, G.A. Role of salt intake in prevention of cardiovascular disease: Controversies and challenges. Nat. Rev. Cardiol. 2018, 15, 371–377. [Google Scholar] [CrossRef]
- Santos, E.; Bhlns, M.; Ferriani, A.; Teixeira, S. Flavonoids: Classification, Biosynthesis and Chemical Ecology; Flavonoids—From Biosynthesis to Human Health; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Iwashina, T. Flavonoid properties of five families newly incorporated into the order Caryophyllales. Bull. Natl. Mus. Nat. Sci. 2013, 39, 25–51. [Google Scholar]
- Iinuma, M.; Tanaka, T.; Hamada, K.; Mizuno, M.; Asai, F.; Reher, G.; Kraus, L. Revised structure of neoflavone in Coutarea hexandra. Phytochemestry 1987, 26, 3096–3097. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Luo, M.; Wei, S. The bioprotective effects of polyphenols on metabolic syndrome against oxidative stress: Evidences and perspectives. Oxidative Med. Cell. Longev. 2019, 2019, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hoensch, H.P.; Oertel, R. The value of flavonoids for the human nutrition: Short review and perspectives. Clin. Nutr. Exp. 2015, 3, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Siow, R.C.; Mann, G.E. Dietary isoflavones and vascular protection: Activation of cellular antioxidant defenses by SERMs or hormesis? Mol. Asp. Med. 2010, 31, 468–477. [Google Scholar] [CrossRef]
- Corcoran, M.P.; McKay, D.L.; Blumberg, J.B. Flavonoid basics: Chemistry, sources, mechanisms of action, and safety. J. Nutr. Gerontol. Geriatr. 2012, 31, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, A.; Szostak-Węgierek, D. Flavonoids–food sources, health benefits, and mechanisms involved. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–27. [Google Scholar]
- Mérillon, J.-M.; Ramawat, K.G. Bioactive Molecules in Food, 1st ed.; Springer Nature: Cham, Switzerland, 2019; pp. 230–256. [Google Scholar]
- Janabi, A.H.W.; Kamboh, A.A.; Saeed, M.; Xiaoyu, L.; BiBi, J.; Majeed, F.; Naveed, M.; Mughal, M.J.; Korejo, N.A.; Kamboh, R. Flavonoid-rich foods (FRF): A promising nutraceutical approach against lifespan-shortening diseases. Iran. J. Basic Med. Sci. 2020, 23, 140. [Google Scholar]
- Kackov, S.; Simundic, A.-M.; Nikolac, N.; Celap, I.; Dukic, L.; Ruzic, D.; Bilusic, M. The effect of high-calorie meal consumption on oxidative stress and endothelial dysfunction in healthy male adults. Physiol. Res. 2013, 62, 643. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Bandy, B. Mechanisms of flavonoid protection against myocardial ischemia–reperfusion injury. J. Mol. Cell. Cardiol. 2009, 46, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H. Flavonoids, dairy foods, and cardiovascular and metabolic health: A review of emerging biologic pathways. Circ. Res. 2018, 122, 369–384. [Google Scholar] [CrossRef]
- Goya, L.; Martín, M.Á.; Sarriá, B.; Ramos, S.; Mateos, R.; Bravo, L. Effect of cocoa and its flavonoids on biomarkers of inflammation: Studies of cell culture, animals and humans. Nutrients 2016, 8, 212. [Google Scholar] [CrossRef]
- Hollman, P.C.; Katan, M.B. Bioavailability and health effects of dietary flavonols in man. In Diversification in Toxicology: Man and Environment, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 237–248. [Google Scholar]
- di Gesso, J.L.; Kerr, J.S.; Zhang, Q.; Raheem, S.; Yalamanchili, S.K.; O’Hagan, D.; Kay, C.D.; O’Connell, M.A. Flavonoid metabolites reduce tumor necrosis factor-α secretion to a greater extent than their precursor compounds in human THP-1 monocytes. Mol. Nutr. Food Res. 2015, 59, 1143–1154. [Google Scholar] [CrossRef] [Green Version]
- Warner, E.F.; Zhang, Q.; Raheem, K.S.; O’Hagan, D.; O’Connell, M.A.; Kay, C.D. Common phenolic metabolites of flavonoids, but not their unmetabolized precursors, reduce the secretion of vascular cellular adhesion molecules by human endothelial cells. J. Nutr. 2016, 146, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Millán, E.; Ramos, S.; Alvarez, C.; Bravo, L.; Goya, L.; Martín, M.Á. Microbial phenolic metabolites improve glucose-stimulated insulin secretion and protect pancreatic beta cells against tert-butyl hydroperoxide-induced toxicity via ERKs and PKC pathways. Food Chem. Toxicol. 2014, 66, 245–253. [Google Scholar] [CrossRef]
- Amin, H.P.; Czank, C.; Raheem, S.; Zhang, Q.; Botting, N.P.; Cassidy, A.; Kay, C.D. Anthocyanins and their physiologically relevant metabolites alter the expression of IL-6 and VCAM-1 in CD40L and oxidized LDL challenged vascular endothelial cells. Mol. Nutr. Food Res. 2015, 59, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Sun, H.; Zha, W.; Cui, W.; Xu, L.; Min, Q.; Wu, J. Apigenin attenuates adriamycin-induced cardiomyocyte apoptosis via the PI3K/AKT/mTOR pathway. Evid. Based Complement. Alternat. Med. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yang, J.; Hu, J.; Li, X.; Zhang, X.; Li, Z. Apigenin attenuates myocardial ischemia/reperfusion injury via the inactivation of p38 mitogen-activated protein kinase. Mol. Med. Rep. 2015, 12, 6873–6878. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Di, S.; Hu, W.; Feng, Y.; Zhou, Q.; Gong, B.; Tang, X.; Liu, J.; Zhang, W.; Xi, M. A new flavonoid glycoside (APG) isolated from Clematis tangutica attenuates myocardial ischemia/reperfusion injury via activating PKCε signaling. Biochim. Biophys. Acta Mol. Basis Dis. (BBA) Mol. Basis Dis. 2017, 1863, 701–711. [Google Scholar] [CrossRef]
- Sun, Z.; Lu, W.; Lin, N.; Lin, H.; Zhang, J.; Ni, T.; Meng, L.; Zhang, C.; Guo, H. Dihydromyricetin alleviates doxorubicin-induced cardiotoxicity by inhibiting NLRP3 inflammasome through activation of SIRT1. Biochem. Pharmacol. 2020, 175, 113888. [Google Scholar] [CrossRef]
- Patel, R.V.; Mistry, B.M.; Shinde, S.K.; Syed, R.; Singh, V.; Shin, H.-S. Therapeutic potential of quercetin as a cardiovascular agent. Eur. J. Med. Chem. 2018, 155, 889–904. [Google Scholar] [CrossRef]
- Ni, T.; Lin, N.; Huang, X.; Lu, W.; Sun, Z.; Zhang, J.; Lin, H.; Chi, J.; Guo, H. Icariin ameliorates diabetic cardiomyopathy through Apelin/Sirt3 Signalling to improve mitochondrial dysfunction. Front. Pharmacol. 2020, 11, 256. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, P.; Zhang, X.; Ma, Y.; Li, W.; Chen, J.-M.; Guo, H.-M.; Bucala, R.; Zhuang, J.; Li, J. Natural antioxidant-isoliquiritigenin ameliorates contractile dysfunction of hypoxic cardiomyocytes via AMPK signaling pathway. Mediators Inflamm. 2013, 2013. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, X.; Chen, L.; Zhou, X.; Zheng, G.; Zhang, H.; Huang, W.; Cai, J. Anti-fibrosis effect of scutellarin via inhibition of endothelial–mesenchymal transition on isoprenaline-induced myocardial fibrosis in rats. Molecules 2014, 19, 15611–15623. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Shibu, M.A.; Fan, M.-J.; Chen, M.-C.; Viswanadha, V.P.; Lin, Y.-L.; Lai, C.-H.; Lin, K.-H.; Ho, T.-J.; Kuo, W.-W. Purple rice anthocyanin extract protects cardiac function in STZ-induced diabetes rat hearts by inhibiting cardiac hypertrophy and fibrosis. J. Nutr. Biochem. 2016, 31, 98–105. [Google Scholar] [CrossRef]
- Al Numair, K.S.; Chandramohan, G.; Alsaif, M.A.; Baskar, A.A. Protective effect of morin on cardiac mitochondrial function during isoproterenol-induced myocardial infarction in male Wistar rats. Redox Rep. 2012, 17, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Kuzu, M.; Kandemir, F.M.; Yildirim, S.; Kucukler, S.; Caglayan, C.; Turk, E. Morin attenuates doxorubicin-induced heart and brain damage by reducing oxidative stress, inflammation and apoptosis. Biomed. Pharmacother. 2018, 106, 443–453. [Google Scholar] [CrossRef]
- Garg, S.; Malhotra, R.K.; Khan, S.I.; Sarkar, S.; Susrutha, P.; Singh, V.; Goyal, S.; Nag, T.C.; Ray, R.; Bhatia, J. Fisetin attenuates isoproterenol-induced cardiac ischemic injury in vivo by suppressing RAGE/NF-κB mediated oxidative stress, apoptosis and inflammation. Phytomedicine 2019, 56, 147–155. [Google Scholar] [CrossRef]
- Sali, V.K.; Vasanthi, H.R. Protective effect of rutin isolated from Spermococe hispida against cobalt chloride-induced hypoxic injury in H9c2 cells by inhibiting oxidative stress and inducing apoptosis. Phytomed. 2018, 51, 196–204. [Google Scholar]
- Wu, W.Y.; Cui, Y.K.; Hong, Y.X.; Li, Y.D.; Wu, Y.; Li, G.; Li, G.R.; Wang, Y. Doxorubicin cardiomyopathy is ameliorated by acacetin via Sirt1-mediated activation of AMPK/Nrf2 signal molecules. J. Cell. Mol. Med. 2020, 24, 12141–12153. [Google Scholar] [CrossRef]
- Maneesai, P.; Bunbupha, S.; Potue, P.; Berkban, T.; Kukongviriyapan, U.; Kukongviriyapan, V.; Prachaney, P.; Pakdeechote, P. Hesperidin prevents nitric oxide deficiency-induced cardiovascular remodeling in rats via suppressing TGF-β1 and MMPs protein expression. Nutrients 2018, 10, 1549. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ma, C.; Liu, C.; Wei, F. Luteolin attenuates doxorubicin-induced cardiotoxicity by modulating the PHLPP1/AKT/Bcl-2 signalling pathway. PeerJ 2020, 8, e8845. [Google Scholar] [CrossRef]
- Xiao, C.; Xia, M.-L.; Wang, J.; Zhou, X.-R.; Lou, Y.-Y.; Tang, L.-H.; Zhang, F.-J.; Yang, J.-T.; Qian, L.-B. Luteolin attenuates cardiac ischemia/reperfusion injury in diabetic rats by modulating Nrf2 antioxidative function. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Cui, G.; Luk, S.C.W.; Li, R.A.; Chan, K.K.K.; Lei, S.W.; Wang, L.; Shen, H.; Leung, G.P.H.; Lee, S.M.Y. Cytoprotection of baicalein against oxidative stress-induced cardiomyocytes injury through the Nrf2/Keap1 pathway. J. Cardiovasc. Pharmacol. 2015, 65, 39–46. [Google Scholar] [CrossRef]
- Li, Q.; Yu, Z.; Xiao, D.; Wang, Y.; Zhao, L.; An, Y.; Gao, Y. Baicalein inhibits mitochondrial apoptosis induced by oxidative stress in cardiomyocytes by stabilizing MARCH5 expression. J. Cell. Mol. Med. 2020, 24, 2040–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, L.; Gao, J.; Lin, H.; Qu, Y.; Shang, C.; Wang, Y.; Lu, Y.; Cui, X. Regulatory Mechanisms of Baicalin in Cardiovascular Diseases: A Review. Front. Pharmacol. 2020, 11, 1601. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Han, J.; Ren, H.; Yang, W.; Zhang, X.; Zheng, Q.; Wang, D. Cardioprotective Effects of Astragalin against Myocardial Ischemia/Reperfusion Injury in Isolated Rat Heart. Oxidative Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, X.; Lv, Z.-Y.; Yin, M.-J.; Chen, J.; Wang, J.; Wu, Q.-N. The Protective Effect of Cyanidin-3-Glucoside on Myocardial Ischemia-Reperfusion Injury through Ferroptosis. Oxidative Med. Cell. Longev. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Wang, C.; Li, X.; Liu, Z.; Han, M.-L.; Hou, Y.-L.; Guo, C.-L. The effect and mechanism of hyperoside on high glucose-induced oxidative stress injury of myocardial cells. Sichuan Da Xue Xue Bao Yi Xue Ban 2018, 49, 518–523. [Google Scholar]
- Liu, Z.; Song, X.D.; Xin, Y.; Wang, X.J.; Yu, H.; Bai, Y.Y.; Liu, J.H.; Zhang, C.N.; Hui, R.T. Protective effect of chrysoeriol against doxorubicin-induced cardiotoxicity in vitro. Chinese Med. J. 2009, 122, 2652–2656. [Google Scholar]
- Liu, L.; Wu, Y.; Huang, X. Orientin protects myocardial cells against hypoxia-reoxygenation injury through induction of autophagy. Eur. J. Pharmacol. 2016, 776, 90–98. [Google Scholar] [CrossRef]
- Dong, L.-Y.; Li, S.; Zhen, Y.-L.; Wang, Y.-N.; Shao, X.; Luo, Z.-G. Cardioprotection of vitexin on myocardial ischemia/reperfusion injury in rat via regulating inflammatory cytokines and MAPK pathway. Am. J. Chinese Med. 2013, 41, 1251–1266. [Google Scholar] [CrossRef]
- Feng, H.; Cao, J.; Zhang, G.; Wang, Y. Kaempferol attenuates cardiac hypertrophy via regulation of ASK1/MAPK signaling pathway and oxidative stress. Planta Med. 2017, 83, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Pengnet, S.; Prommaouan, S.; Sumarithum, P.; Malakul, W. Naringin reverses high-cholesterol diet-induced vascular dysfunction and oxidative stress in rats via regulating LOX-1 and NADPH oxidase subunit expression. BioMed Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ramprasath, T.; Senthamizharasi, M.; Vasudevan, V.; Sasikumar, S.; Yuvaraj, S.; Selvam, G.S. Naringenin confers protection against oxidative stress through upregulation of Nrf2 target genes in cardiomyoblast cells. J. Physiol. Biochem. 2014, 70, 407–415. [Google Scholar] [CrossRef]
- Tian, L.; Cao, W.; Yue, R.; Yuan, Y.; Guo, X.; Qin, D.; Xing, J.; Wang, X. Pretreatment with Tilianin improves mitochondrial energy metabolism and oxidative stress in rats with myocardial ischemia/reperfusion injury via AMPK/SIRT1/PGC-1 alpha signaling pathway. J. Pharmacol. Sci. 2019, 139, 352–360. [Google Scholar] [CrossRef]
- Gu, M.; He, P.; Lyu, C.; Liu, X.; Xu, Y.; Cheng, S.; Gu, Y.; Jia, Y. Spinosin and 6′’’-Feruloylspinosin protect the heart against acute myocardial ischemia and reperfusion in rats. Mol. Med. Rep. 2019, 20, 4253–4261. [Google Scholar] [CrossRef] [Green Version]
- Scarabelli, T.M.; Mariotto, S.; Abdel-Azeim, S.; Shoji, K.; Darra, E.; Stephanou, A.; Chen-Scarabelli, C.; Marechal, J.D.; Knight, R.; Ciampa, A. Targeting STAT1 by myricetin and delphinidin provides efficient protection of the heart from ischemia/reperfusion-induced injury. FEBS Lett. 2009, 583, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.-S.; Lin, Y.-M.; Kuo, W.-W.; Pan, L.-F.; Yeh, Y.-L.; Li, Y.-H.; Kuo, C.-H.; Chen, R.-J.; Padma, V.V.; Chen, T.-S. Suppression of isoproterenol-induced apoptosis in H9c2 cardiomyoblast cells by daidzein through activation of Akt. Chin. J. Physiol. 2016, 59, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Samuel, V.P.; Wu, Y.; Dang, M.; Lin, Y.; Sriramaneni, R.; Sah, S.K.; Chinnaboina, G.K.; Zhang, G. Nrf2/HO-1 mediated protective activity of genistein against doxorubicin-induced cardiac toxicity. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 143–152. [Google Scholar] [CrossRef]
- Wei, H.; Li, H.; Wan, S.-P.; Zeng, Q.-T.; Cheng, L.-X.; Jiang, L.-L.; Peng, Y.-D. Cardioprotective effects of Malvidin against isoproterenol-induced myocardial infarction in rats: A mechanistic study. Med. Sci. Mon. Int. Med. J. Exp. Clin. Res. 2017, 23, 2007. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Yang, C.; Shao, L.; Zhu, H.; Wang, Y.; Huang, X.; Wang, S.; Hong, L. Targeting NOX 4 by petunidin improves anoxia/reoxygenation-induced myocardium injury. Eur. J. Pharmacol. 2020, 888, 173414. [Google Scholar] [CrossRef]
- Shabalala, S.C.; Dludla, P.V.; Muller, C.J.; Nxele, X.; Kappo, A.P.; Louw, J.; Johnson, R. Aspalathin ameliorates doxorubicin-induced oxidative stress in H9c2 cardiomyoblasts. Toxicol. Vitro 2019, 55, 134–139. [Google Scholar] [CrossRef]
- Senthamizhselvan, O.; Manivannan, J.; Silambarasan, T.; Raja, B. Diosmin pretreatment improves cardiac function and suppresses oxidative stress in rat heart after ischemia/reperfusion. Eur. J. Pharmacol. 2014, 736, 131–137. [Google Scholar] [CrossRef]
- Bei, W.; Jing, L.; Chen, N. Cardio protective role of wogonin loaded nanoparticle against isoproterenol induced myocardial infarction by moderating oxidative stress and inflammation. Colloids Sur. B Biointerfaces 2020, 185, 110635. [Google Scholar] [CrossRef]
- Zhang, E.; Yang, H.; Li, M.; Ding, M. A possible underlying mechanism behind the cardioprotective efficacy of tangeretin on isoproterenol triggered cardiotoxicity via modulating PI3K/Akt signaling pathway in a rat model. J. Food Biochem. 2020, 44, e13368. [Google Scholar] [CrossRef]
- Sahu, B.D.; Anubolu, H.; Koneru, M.; Kumar, J.M.; Kuncha, M.; Rachamalla, S.S.; Sistla, R. Cardioprotective effect of embelin on isoproterenol-induced myocardial injury in rats: Possible involvement of mitochondrial dysfunction and apoptosis. Life Sci. 2014, 107, 59–67. [Google Scholar] [CrossRef]
- Lalitha, G.; Poornima, P.; Archanah, A.; Padma, V.V. Protective effect of neferine against isoproterenol-induced cardiac toxicity. Carduiovasc. Toxicol. 2013, 13, 168–179. [Google Scholar] [CrossRef]
- Suchal, K.; Malik, S.; Gamad, N.; Malhotra, R.K.; Goyal, S.N.; Ojha, S.; Kumari, S.; Bhatia, J.; Arya, D.S. Mangiferin protect myocardial insults through modulation of MAPK/TGF-β pathways. Eur. J. Pharmacol. 2016, 776, 34–43. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, J.; Liu, W.; Liu, N.; Fu, X.; Kwan, H.; Liu, S.; Liu, B.; Zhang, S.; Yu, Z. Calycosin inhibits oxidative stress-induced cardiomyocyte apoptosis via activating estrogen receptor-α/β. Bioorg. Med. Chem. Lett. 2016, 26, 181–185. [Google Scholar] [CrossRef]
- Yuan, X.; Niu, H.T.; Wang, P.L.; Lu, J.; Zhao, H.; Liu, S.H.; Zheng, Q.S.; Li, C.G. Cardioprotective effect of licochalcone D against myocardial ischemia/reperfusion injury in langendorff-perfused rat hearts. PLoS ONE 2015, 10, e0128375. [Google Scholar] [CrossRef]
- Kim, D.-E.; Kim, B.; Shin, H.-S.; Kwon, H.J.; Park, E.-S. The protective effect of hispidin against hydrogen peroxide-induced apoptosis in H9c2 cardiomyoblast cells through Akt/GSK-3β and ERK1/2 signaling pathway. Exp. Cell Res. 2014, 327, 264–275. [Google Scholar] [CrossRef]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Ciumărnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, Ș.C.; Răchișan, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.-G.; Para, I. The Effects of Flavonoids in Cardiovascular Diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; Van Nood, E.; Van Hoorn, D.E.; Boelens, P.G.; Van Norren, K.; Van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Acton, J.P.; Cocksedge, S.P.; Davies, K.A.B.; Bailey, S.J. The effect of dietary phytochemicals on nuclear factor erythroid 2-related factor 2 (Nrf2) activation: A systematic review of human intervention trials. Mol. Biol. Rep. 2021, 48, 1745–1761. [Google Scholar] [CrossRef] [PubMed]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Alexander, M.Y.; White, S.J. The Role of Nrf2 in Cardiovascular Function and Disease. Oxidative Med. Cell. Longev. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed]
- L Suraweera, T.; Rupasinghe, H.; Dellaire, G.; Xu, Z. Regulation of Nrf2/ARE Pathway by Dietary Flavonoids: A Friend or Foe for Cancer Management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef]
- Sugamura, K.; Keaney, J.F., Jr. Reactive oxygen species in cardiovascular disease. Free Radic. Biol. Med. 2011, 51, 978–992. [Google Scholar] [CrossRef] [Green Version]
- Alfaddagh, A.; Martin, S.S.; Leucker, T.M.; Michos, E.D.; Blaha, M.J.; Lowenstein, C.J.; Jones, S.R.; Toth, P.P. Inflammation and cardiovascular disease: From mechanisms to therapeutics. Am. J. Prev. Cardiol. 2020, 4, 100130. [Google Scholar] [CrossRef]
- Choy, K.W.; Murugan, D.; Leong, X.-F.; Abas, R.; Alias, A.; Mustafa, M.R. Flavonoids as natural anti-inflammatory agents targeting nuclear factor-kappa B (NFκB) signaling in cardiovascular diseases: A mini review. Front. Pharmacol. 2019, 10, 1295. [Google Scholar] [CrossRef] [Green Version]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and Mitochondria: Activation of Cytoprotective Pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef]
- Crow, M.T.; Mani, K.; Nam, Y.-J.; Kitsis, R.N. The mitochondrial death pathway and cardiac myocyte apoptosis. Circ. Res. 2004, 95, 957–970. [Google Scholar] [CrossRef] [Green Version]
- Testai, L. Flavonoids and mitochondrial pharmacology: A new paradigm for cardioprotection. Life Sci. 2015, 135, 68–76. [Google Scholar] [CrossRef]
- Montero, M.; Lobatón, C.D.; Hernández-Sanmiguel, E.; Santodomingo, J.; Vay, L.; Moreno, A.; Alvarez, J. Direct activation of the mitochondrial calcium uniporter by natural plant flavonoids. Biochem.J. 2004, 384, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Bednarczyk, P.; Kicinska, A.; Jarmuszkiewicz, W.; Debowska, R.; Szewczyk, A. Flavonoids as natural modulators of mitochondrial potassium channel. Biophys. J. 2017, 112, 405a–406a. [Google Scholar] [CrossRef] [Green Version]
- Liobikas, J.; Skemiene, K.; Trumbeckaite, S.; Borutaite, V. Anthocyanins in cardioprotection: A path through mitochondria. Pharmacol. Res. 2016, 113, 808–815. [Google Scholar] [CrossRef]
- Lagoa, R.; Graziani, I.; Lopez-Sanchez, C.; Garcia-Martinez, V.; Gutierrez-Merino, C. Complex I and cytochrome c are molecular targets of flavonoids that inhibit hydrogen peroxide production by mitochondria. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 1562–1572. [Google Scholar] [CrossRef] [Green Version]
- Serreli, G.; Deiana, M. In vivo formed metabolites of polyphenols and their biological efficacy. Food Funct. 2019, 10, 6999–7021. [Google Scholar] [CrossRef]
- Yang, B.; Liu, H.; Yang, J.; Gupta, V.K.; Jiang, Y. New insights on bioactivities and biosynthesis of flavonoid glycosides. Trends Food Sci.Technol. 2018, 79, 116–124. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The bioavailability, transport, and bioactivity of dietary flavonoids: A review from a historical perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef] [Green Version]
- Kamiloglu, S.; Tomas, M.; Ozdal, T.; Capanoglu, E. Effect of food matrix on the content and bioavailability of flavonoids. Trends Food Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Chen, Z.; Zheng, S.; Li, L.; Jiang, H. Metabolism of flavonoids in human: A comprehensive review. Curr. Drug Metab. 2014, 15, 48–61. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, H.-H. Effects of the flavonoids on cytochrome P-450 CYP1, 2E1, 3A4 and 19. Yao Xue Xue Bao 2007, 42, 8–12. [Google Scholar]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [Green Version]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [Green Version]
- Galati, G.; O’brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef]
- Sak, K. Cytotoxicity of dietary flavonoids on different human cancer types. Pharmacogn. Rev. 2014, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Yáñez, J.A.; Chemuturi, N.V.; Womble, S.W.; Sayre, C.L.; Davies, N.M. Flavonoids and drug interactions. In Flavonoid Pharmacokinetics: Methods of Analysis, Pre-Clinical and Clinical Pharmacokinetics, Safety, and Toxicology, 1st ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2012; pp. 281–319. [Google Scholar]
- Rosenkranz, B.; Fasinu, P.; Bouic, P. An overview of the evidence and mechanisms of herb–drug interactions. Front. Pharmacol. 2012, 3, 69. [Google Scholar]
- Branquinho Andrade, P.; Grosso, C.; Valentao, P.; Bernardo, J. Flavonoids in neurodegeneration: Limitations and strategies to cross CNS barriers. Curr. Med. Chem. 2016, 23, 4151–4174. [Google Scholar] [CrossRef]
- Castangia, I.; Nácher, A.; Caddeo, C.; Merino, V.; Díez-Sales, O.; Catalán-Latorre, A.; Fernàndez-Busquets, X.; Fadda, A.M.; Manconi, M. Therapeutic efficacy of quercetin enzyme-responsive nanovesicles for the treatment of experimental colitis in rats. Acta Biomater. 2015, 13, 216–227. [Google Scholar] [CrossRef]
- Colone, M.; Calcabrini, A.; Stringaro, A. Drug Delivery Systems of Natural Products in Oncology. Molecules 2020, 25, 4560. [Google Scholar] [CrossRef] [PubMed]
- Yeh, M.I.; Huang, H.C.; Liaw, J.H.; Huang, M.C.; Wu, T.H.; Huang, K.F.; Hsu, F.L. Ethosomes in hair dye products as carriers of the major compounds of black tea extracts. Int. J. Dermatol. 2013, 52, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Li, Y.; McClements, D.J.; Xiao, H. Nanoemulsion-and emulsion-based delivery systems for curcumin: Encapsulation and release properties. Food Chem. 2012, 132, 799–807. [Google Scholar] [CrossRef]
- Thilakarathna, S.H.; Rupasinghe, H. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharmaceut 2019, 570, 118642. [Google Scholar] [CrossRef]
- Bisol, Â.; de Campos, P.S.; Lamers, M.L. Flavonoids as anticancer therapies: A systematic review of clinical trials. Phytother. Res. 2020, 34, 568–582. [Google Scholar] [CrossRef]
- Amin, M.U.; Khurram, M.; Khattak, B.; Khan, J. Antibiotic additive and synergistic action of rutin, morin and quercetin against methicillin resistant Staphylococcus aureus. BMC Compl. Alternative Med. 2015, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- White, J.R. Apple trees to sodium glucose co-transporter inhibitors: A review of SGLT2 inhibition. Clin. Diabetes 2010, 28, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Lalani, S.; Poh, C.L. Flavonoids as antiviral agents for Enterovirus A71 (EV-A71). Viruses 2020, 12, 184. [Google Scholar] [CrossRef] [Green Version]
- Liskova, A.; Samec, M.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Al-Ishaq, R.K.; Abotaleb, M.; Nosal, V.; Kajo, K.; Ashrafizadeh, M. Flavonoids against the SARS-CoV-2 induced inflammatory storm. Biomed. Pharmacother. 2021, 138, 111430. [Google Scholar] [CrossRef]
- Rudrapal, M.; Chetia, D. Plant flavonoids as potential source of future antimalarial leads. Sys. Rev. Pharm. 2017, 8, 13–18. [Google Scholar] [CrossRef]
- Rasmi, Y.; Li, X.; Khan, J.; Ozer, T.; Choi, J.R. Emerging point-of-care biosensors for rapid diagnosis of COVID-19: Current progress, challenges, and future prospects. Anal. Bioanal. Chem. 2021, 1–23. [Google Scholar] [CrossRef]
- Bhat, E.A. SARS-CoV-2: Insight in genome structure, pathogenesis and viral receptor binding analysis–An Updated Review. Int. Immunopharmacol. 2021, 95, 107493. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Major Flavonoids | Major Source | Health Benefits |
|---|---|---|---|
| Flavonols | Isorhamnetin Kaempferol Myricetin Quercetin | Onions, broccoli, tea, apple, blueberries | Regulates systolic blood pressure, glycemic levels, and BMI. |
| Flavones | Apigenin Luteolin | Parsley, celery, chamomile tea, fenugreek, onion, garlic, pepper, citrus fruits | Regulates blood glucose levels. |
| Flavanones | Eriodictyol Hesperetin Naringenin | Citrus fruits, mint, tomatoes. | Lowers risk of ischaemic stroke. |
| Flavanols | Catechins Epicatechins | Apricots, cocoa, chocolates, red grapes, red wine, tea | Reduces mean arterial pressure. Improves insulin resistance and LDL-C, HDL-C levels. |
| Procyanidins | Theaflavins Thearubigins | Cocoa, apples, grapes, red wine, chocolates | Regulates blood pressure. |
| Anthocyanidins | Cyanidin Delphinidin Malvidin Pelargonidin Peonidin Petunidin | Berries, red wine, red cabbage, bright colored fruits, cherries, cranberries | Lowers risk of Myocardial infarctions. |
| Isoflavones | Daidzein Genistein Glycitein | Soyabean, dairy products, egg, meat | Beneficial for T2DM. |
| Flavonoids | Oxidative Stress Model | Molecular Mechanism | Reference (s) |
|---|---|---|---|
| Apigenin | Myocardial ischemia-reperfusion injury in h9C2 cardiomyocytes; adriamycin-induced cardiotoxicity in Kunming mice | ↑PI3K/AKT/mTOR pathway | [37,38] |
| Apigenin-7-O-b-D-(-6′′-p-coumaroyl)-glucopyranoside | Primary neonatal cardiomyocyte (C57/6J) ischemic reperfusion model in vitro | ↑PKCe translocation signaling ↑Nrf2/HO-1 pathway ↓NF-kB signaling Pathway | [39] |
| Dihydromyricetin | Doxorubicin-induced cardiotoxicity | ↑SIRT1 ↓NLRP3 inflammosome | [40] |
| Quercetin | Isoproterenol-induced cardiac fibrosis | ↑Nrf2-HO; ↓LDL receptor expression; ROS scavenger | [41] |
| Icarin | High glucose- and adenovirus-induced cardiomyopathy in neonatal C57 mice | ↑Apelin/SIRT3 | [42] |
| Isoliquiritigenin | Hypoxia-induced contractile dysfunction in cardiomyocytes | ↑AMPK and ERK signaling pathways; ROS scavenger | [43] |
| Scutellarin | Isoproterenol-induced myocardial infarction in SD rats | ↓α-SMA ↑CD31, Jagged1, Notch 1, and Hes1 | [44] |
| Cyanidin-3-glucoside | Wistar rats induced by STZ | ↑TIMP-1 ↓MMP-9,.TGF-b, p-MEK1/2, CTGF, P-ERK1/2, FGF2 | [45] |
| Morin | Isoproterenol-induced myocardial infarction; doxorubicin-induced cardiac fibrosis | Restored the mitochondrial function and improvement of mitochondrial antioxidant enzymes; ↓myocardial; Apoptosis; ↑Bcl-2 | [46,47] |
| Fisetin | Isoproterenol-induced cardiac ischemia | ↓RAGE and NF-κB; ↓Bax, caspase-3, cytochrome-c; ↑Bcl-2; ↓Myocardial apoptosis | [48] |
| Rutin | Cobalt chloride-induced hypoxic injury in H9c2 cells | Modulation of Akt, p-Akt, p38 and p-p38; ↓of HIF-1α, BAX and caspase | [49] |
| Acacetin | Doxorubicin-induced cardiomyopathy | ↑Sirt1/pAMPK pathway ↑AMPK/Nrf2 signal pathway | [50] |
| Hesperidin | Nitric oxide deficiency-induced cardiovascular remodeling | ↓TNF-R1 and TGF- β1 protein expression; ↓MMP-2 and MMP-9 | [51] |
| Luteolin | Doxorubicin-induced cardiotoxicity | ↑AKT/Bcl-2 signaling pathway; ↑Nrf2/HO-1 pathway; ↑eNOS/Nrf2 signaling pathway | [52,53] |
| Baicalein | t-BHP-induced oxidative stress; H2O2 and ischemia/reperfusion (I/R) stress | ↑Nrf2/Keap1 pathway; ↓KLF4-MARCH5-Drp1 pathway | [54,55] |
| Baicalin | Hypoxia-induced oxidative stress in cardiomyocytes; Angiotensin-II-induced endothelial dysfunction | ↑Nrf2/HO-1 signaling pathway; ↓NF-kB signaling pathway; ↓iNOS protein expression | [56,57] |
| Astragallin | Myocardial ischemia/reperfusion (I/R) injury in isolated rat heart | ↓ROS; ↓ Inflammation; ↓Myocardial apoptosis; ↑Bcl-2 | [58] |
| Cyanidin-3-O-glucoside | Myocardial ischemia-reperfusion injury in SD rats and H9c2 cells | ↓USP19, Beclin1, NCOA4, and LC3II/LC3I; ↓LC3II/LC3I; ↓TfR1 expression; ↑FTH1 and GPX4; ↓Ferroptosis promoter RSL3 | [59] |
| Hyperoside | High glucose-induced oxidative stress in cardiac cells | ↑ p-AKT/AKT and p-Nrf2/Nrf2; ↓Myocardial apoptosis and levels of ROS and MDA | [60] |
| Chrysoeriol | Doxorubicin-induced toxicity in cardiomyocytes | ↓ROS, MDA; ↑GSH, SOD | [61] |
| Orientin | Myocardial ischemia reperfusion injury | ↑AMPK, Akt and Bcl-2; ↓mTOR and Raptor, Beclin 1 | [62] |
| Vitexin | Myocardial ischemia/reperfusion (I/R) injury | ↓phospho-c-Jun; ↑phospho-ERK; ↓inflammatory cytokines and ↓MAPK pathway. | [63] |
| Kaempferol | Cardiac hypertrophy by aorta banding | ↓ASK1/JNK1/2/p38 signaling pathway; ↓ASK1/MAPK signaling pathways (JNK1/2 and p38) | [64] |
| Naringin | High-cholesterol-diet-induced endothelial dysfunction and oxidative stress in rats | ↓LOX-1, NADPH oxidase subunits (p47phox, Nox2, and Nox4), and iNOS | [65] |
| Naringenin | H2O2-induced oxidative stress in cardiomyocytes | ↓ROS; ↑Nrf2 signaling pathway | [66] |
| Tilianin | Myocardial ischemia/reperfusion injury in rats | ↑AMPK, pAMPK, SIRT1, PGC-1alpha, NRF1, TFAM and FOXO1 proteins | [67] |
| Spinosin | Myocardial ischemia/reperfusion injury in rats | ↓GSK3β; ↑PGC-1α; ↑Nrf2/HO-1 pathway | [68] |
| Myricetin | Myocardial ischemia/reperfusion injury in rats | ↓STAT1 | [69] |
| Delphinidin | Myocardial ischemia/reperfusion injury in rats | ↓STAT1 | [69] |
| Daidzein | Isoproterenol-induced apoptosis in H9c2 cardiomyoblast | ↑Akt activation | [70] |
| Genistein | Doxorubicin-induced cardiotoxicity | ↑Nrf2/HO-1 signaling pathway; ↓DNA damage | [71] |
| Malvidin | Isoproterenol-induced apoptosis in H9c2 cardiomyoblast | ↑Nrf2/HO-1 signaling pathway; ↓NF-κB signaling pathway activation | [72] |
| Petunidin | Myocardial ischemia/reperfusion injury in rats | ↑Bcl-2 protein expression, ↓ NOX4 and Bax expression, ↓cytoplasmic cytochrome c expression; ↓ROS | [73] |
| Aspalathin | Doxorubicin-induced cardiotoxicity in cardiomyocytes | ↓ROS; ↓ Myocardial apoptosis | [74] |
| Diosmin | Myocardial ischemia/reperfusion injury in rats | ↑Bcl-2 expression; ↑antioxidant enzyme activities; ↓LPO | [75] |
| Wogonin | Isoproterenol-induced myocardial infarction | ↑Nrf2/HO-1 signaling pathway; ↓Inflammation | [76] |
| Tangeretin | Isoproterenol-induced myocardial infarction | ↑PI3K/Akt signaling pathway | [77] |
| Embelin | Isoproterenol-induced myocardial injury | ↑Bcl-2; ↓Bax, Cytochrome c, cleaved-caspase-3 & 9 and PARP; | [78] |
| Neferin | Isoproterenol-induced myocardial injury | ↓Inflammation; ↑ Tissue antioxidant status | [79] |
| Mangiferin | Myocardial ischemia/reperfusion injury in rats | ↓Phosphorylation of p38 and JNK, phosphorylation of ERK1/2; ↓TGF-β, ↓MAPK | [80] |
| Calycosin | H2O2-induced oxidative stress in cardiomyocytes | ↓ Apoptosis; ↑ ER/ and Akt | [81] |
| Licochalcone D | Myocardial ischemia/reperfusion (I/R) injury in cardiomyocytes | ↓ Caspase 3 and PARP; ↓ IL-6, NF-kB and p38 MAPK | [82] |
| Hispidin | H2O2-induced oxidative stress in cardiomyocytes | ↓ Apoptosis, ROS, DNA damage, caspase 3 and Bax expression ↑ HO-1, CAT, Bcl-2, Akt/GSK3 and ERK ½ | [83] |
| Drugs | Flavonoid | Species in Which Tested | Change in Bioavailability |
|---|---|---|---|
| Diltiazem (15 mg/kg, oral) | Morin (1.5–7.5 mg/kg, oral) | Rat | 1.4- to 1.8-fold increases |
| Talinolol (10 mg/kg, oral) | Naringin (1–20 mg/kg, oral) | Rat | 1.5- to 3.0-foldincreases |
| Etoposide (6 mg/kg, oral) | Morin (15 mg/kg, oral) | Rat | 1.4-fold increases |
| Digoxin (0.02 mg/kg, oral) | Quercetin (40 mg/kg, oral) | Pig | 1.7-foldincreases |
| Moxidectin (0.2 mg/kg, subcutaneous) | Quercetin (10 mg/kg, subcutaneous) | Sheep | 1.8-fold increases |
| Verapamil (10 mg/kg, oral) | Quercetin (15 mg/kg, oral) | Rabbit | 2-fold increases |
| Paclitaxel (30 mg/kg oral) | Genistein (10 mg/kg, oral) | Rat | 1.5-fold increases |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules 2021, 26, 4021. https://doi.org/10.3390/molecules26134021
Khan J, Deb PK, Priya S, Medina KD, Devi R, Walode SG, Rudrapal M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules. 2021; 26(13):4021. https://doi.org/10.3390/molecules26134021
Chicago/Turabian StyleKhan, Johra, Prashanta Kumar Deb, Somi Priya, Karla Damián Medina, Rajlakshmi Devi, Sanjay G. Walode, and Mithun Rudrapal. 2021. "Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns" Molecules 26, no. 13: 4021. https://doi.org/10.3390/molecules26134021