
Generalizing the Wells–Riley Infection Probability: A Superstatistical Scheme for Indoor Infection Risk Estimation
Abstract
:1. Introduction
2. Model Construction
2.1. Preliminaries
2.2. Airborne Exposure Risk Statistics
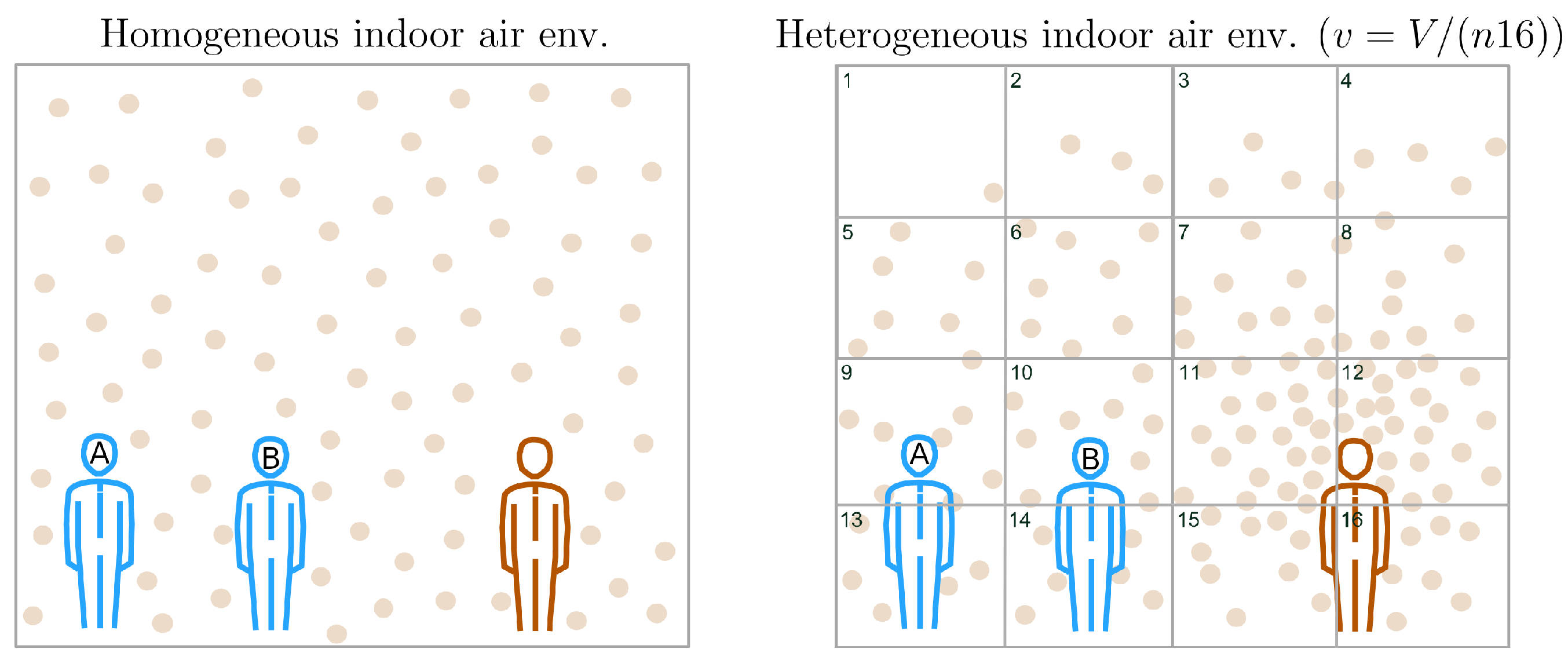
2.2.1. Homogeneous Indoor Air Environment
2.2.2. Heterogeneous Indoor Air Environment
2.3. Infection-Activation Considerations
2.4. Indoor Infection Dynamics
3. Insights Gained from Computational Analysis
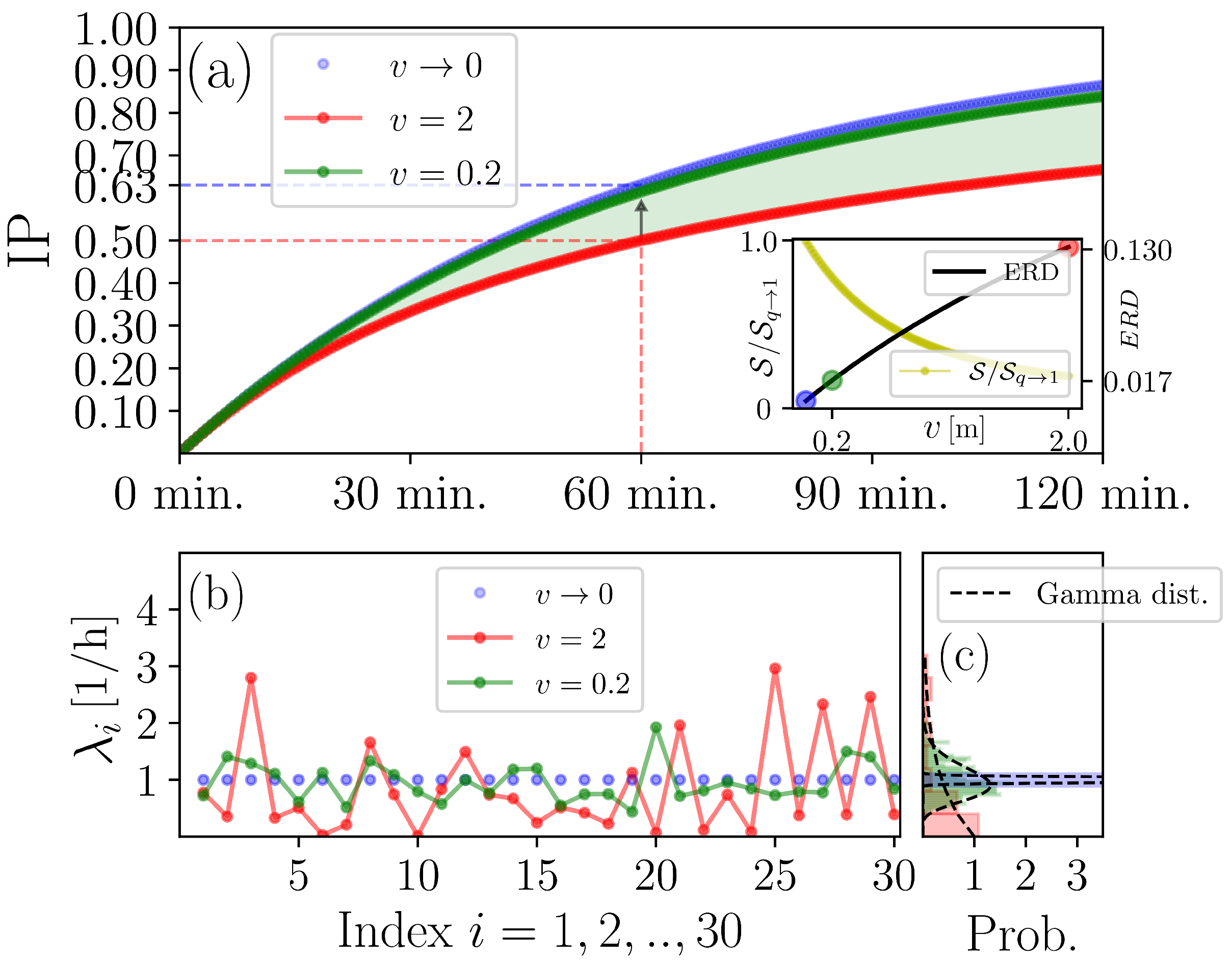
3.1. Scrutinising the Generalised WRIP
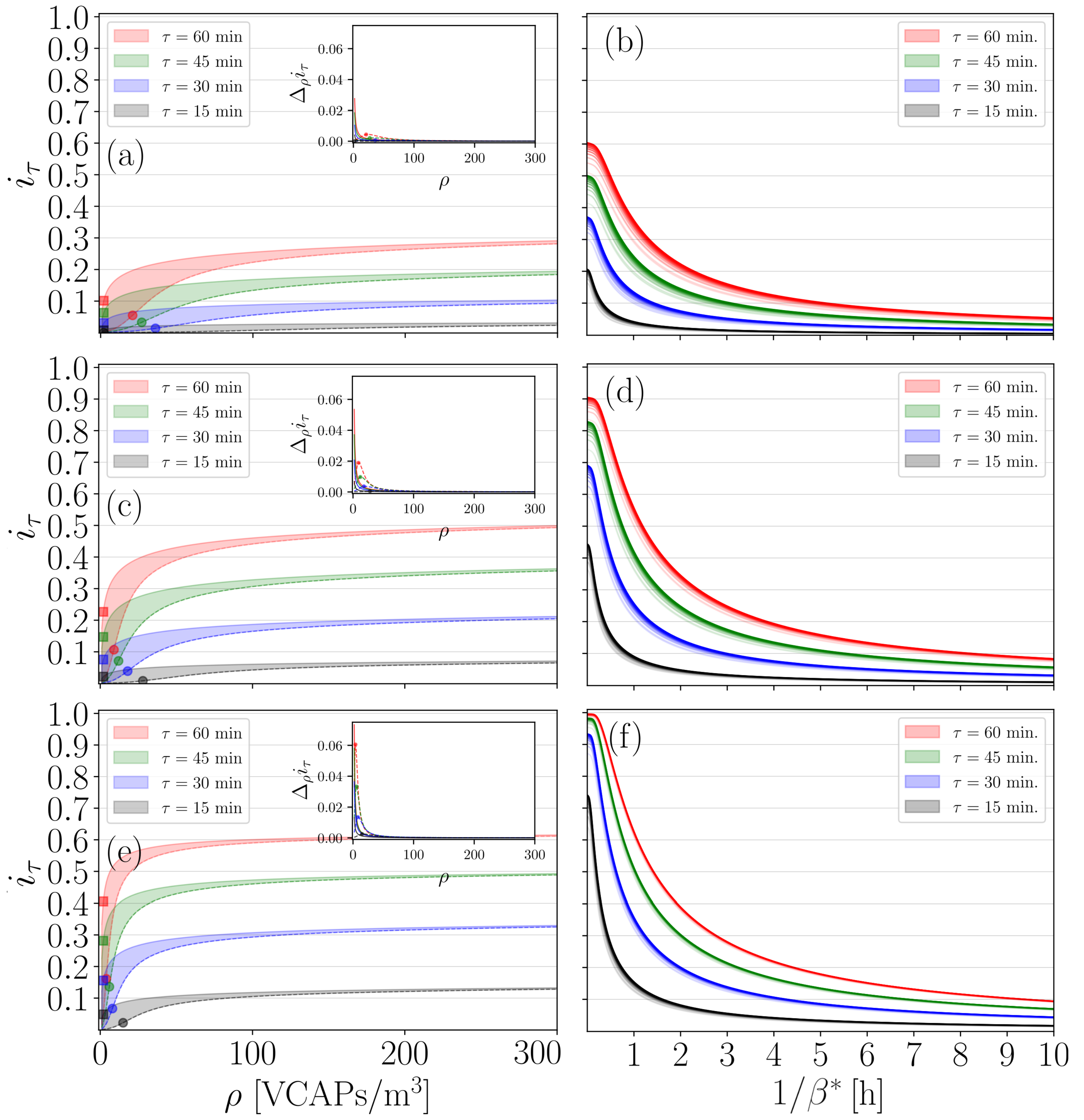
3.2. Refining the IIRE
3.3. Evaluation of the Six-Foot Rule
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A
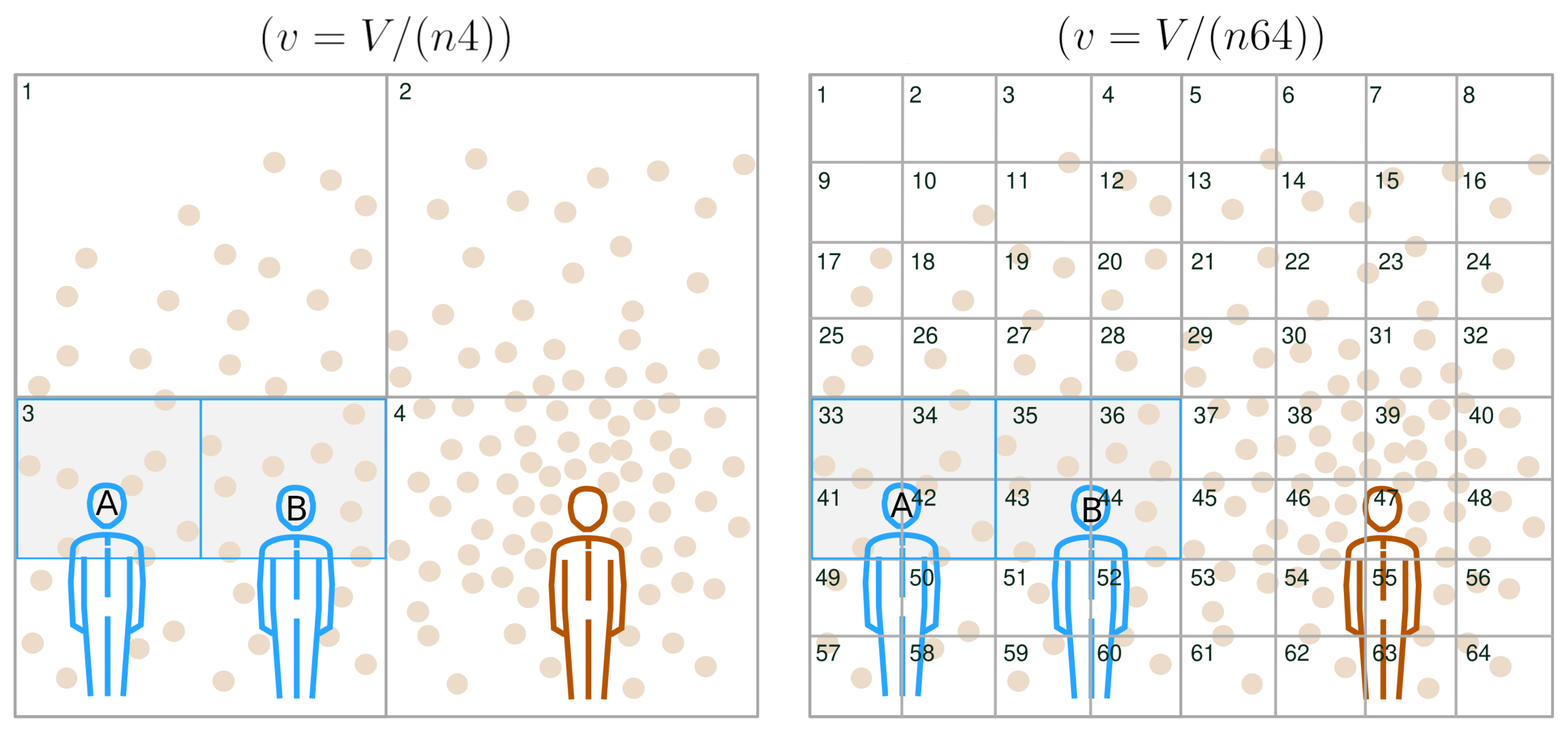
Appendix A.1. Probing the Spatial Configuration of VCAPs
Appendix A.2. Schematic Illustration of the Contaminated-Air-Sharing Scenario

Appendix A.3. Discretisation of the Tsallis Entropic Functional
References
- National Academies of Sciences, Engineering, and Medicine. Airborne Transmission of SARS-CoV-2: Proceedings of a Workshop-in Brief; The National Academies Press: Washington, DC, USA, 2020. [Google Scholar] [CrossRef]
- Prather, K.A.; Marr, L.C.; Schooley, R.T.; McDiarmid, M.A.; Wilson, M.E.; Milton, D.K. Airborne Transmission of SARS-CoV-2. Science 2020, 370, 303–304. [Google Scholar] [PubMed]
- Miller, S.L.; Nazaroff, W.W.; Jimenez, J.L.; Boerstra, A.; Buonanno, G.; Dancer, S.J.; Kurnitski, J.; Marr, L.C.; Morawska, L.; Noakes, C. Transmission of SARS-CoV-2 by Inhalation of Respiratory Aerosol in the Skagit Valley Chorale Superspreading Event. Indoor Air 2021, 31, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, T.; Jimenez, J.L.; Prather, K.A.; Tufekci, Z.; Fisman, D.; Schooley, R. Ten Scientific Reasons in Support of Airborne Transmission of SARS-CoV-2. Lancet 2021, 397, 1603–1605. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.W.; Marr, L.C.; Li, Y.; Dancer, S.J. COVID-19 Has Redefined Airborne Transmission. BMJ 2021, 373, n913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, Y.; Zhang, A.L.; Wang, Y.; Molina, M.J. Identifying airborne transmission as the dominant route for the spread of COVID-19. Proc. Natl. Acad. Sci. USA 2020, 117, 14857–14863. [Google Scholar] [CrossRef]
- Setti, L.; Passarini, F.; De Gennaro, G.; Barbieri, P.; Perrone, M.G.; Borelli, M.; Palmisani, J.; Di Gilio, A.; Piscitelli, P.; Miani, A. Airborne transmission route of COVID-19: Why 2 m/6 feet of inter-personal distance could not be enough. Int. J. Environ. Res. Public Health 2020, 17, 2932. [Google Scholar] [CrossRef] [Green Version]
- Morawska, L.; Milton, D.K. It is time to address airborne transmission of coronavirus disease 2019 (COVID-19). Clin. Infect. Dis. 2020, 71, 2311–2313. [Google Scholar] [CrossRef]
- Peng, Z.; Rojas, A.P.; Kropff, E.; Bahnfleth, W.; Buonanno, G.; Dancer, S.J.; Kurnitski, J.; Li, Y.; Loomans, M.G.; Marr, L.C.; et al. Practical Indicators for Risk of Airborne Transmission in SharedIndoor Environments and Their Application to COVID-19 Outbreaks. Environ. Sci. Technol. 2022, 56, 1125–1137. [Google Scholar] [CrossRef]
- Dick, E.C.; Jennings, L.C.; Mink, K.A.; Wartgow, C.D.; Inborn, S.L. Aerosol Transmission of Rhinovirus Colds. J. Infect. Dis. 1987, 156, 442–448. [Google Scholar] [CrossRef]
- Yu, I.T.; Li, Y.; Wong, T.W.; Tam, W.; Chan, A.T.; Lee, J.H.; Leung, D.Y.; Ho, T. Evidence of Airborne Transmission of the Severe Acute Respiratory Syndrome Virus. N. Engl. J. Med. 2004, 350, 1731–1739. [Google Scholar] [CrossRef] [Green Version]
- Moriarty, L.F.; Plucinski, M.M.; Marston, B.J.; Kurbatova, E.V.; Knust, B.; Murray, E.L.; Pesik, N.; Rose, D.; Fitter, D.; Kobayashi, M.; et al. Public Health Responses to COVID-19 Outbreaks on Cruise Ships—Worldwide, February–March 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 347–352. [Google Scholar] [CrossRef]
- Hamner, L.; Dubbel, P.; Capron, I.; Ross, A.; Jordan, A.; Lee, J.; Lynn, J.; Ball1, A.; Narwal, S.; Russell, S.; et al. High SARS-CoV-2 attack rate following exposure at a choir practice, Skagit County, Washington, March 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 606–610. [Google Scholar] [CrossRef]
- Jayaweera, M.; Perera, H.; Gunawardana, B.; Manatunge, J. Transmission of COVID-19 virus by droplets and aerosols. Environ. Res. 2020, 188, 109819. [Google Scholar] [CrossRef]
- Morawksa, L. Droplet fate in indoor environments, or can we prevent the spread of infection? Indoor Air 2006, 16, 335–347. [Google Scholar]
- Tian, D.; Sun, Y.; Xu, H.; Ye, Q. The emergence and epidemic characteristics of the highly mutated SARS-CoV-2 Omicron variant. J. Med. Virol. 2022, 94, 2376–2383. [Google Scholar] [CrossRef]
- Gaddis, M.D.; Manoranjan, V.S. Modeling the Spread of COVID-19 in Enclosed Spaces. Math. Comput. Appl. 2021, 26, 79. [Google Scholar] [CrossRef]
- Noakes, C.J.; Beggs, C.B.; Sleigh, P.A.; Kerr, K.G. Modelling the transmission of airborne infections in enclosed spaces. Epidemiol. Infect. 2006, 134, 1082–1091. [Google Scholar] [CrossRef]
- Tirnakli, U.; Tsallis, C. Epidemiological Model with Anomalous Kinetics: Early Stages of the COVID-19 Pandemic. Front. Phys. 2020, 8, 613168. [Google Scholar] [CrossRef]
- Vasconcelos, G.L.; Pessoa, N.L.; Silva, N.B.; Macêdo, A.M.; Brum, A.A.; Ospina, R.; Tirnakli, U. Multiple waves of COVID-19: A pathway model approach. Nonlinear Dyn. 2023, 111, 6855–6872. [Google Scholar] [CrossRef]
- Dhawan, S.; Biswas, P. Aerosol Dynamics Model for Estimating the Risk from Short-Range Airborne Transmission and Inhalation of Expiratory Droplets of SARS-CoV-2. Environ. Sci. Technol. 2021, 55, 8987–8999. [Google Scholar] [CrossRef]
- Bazant, M.Z.; Bush, J.W.M. A guideline to limit indoor airborne transmission of COVID-19. Proc. Natl. Acad. Sci. USA 2021, 118, 17. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Niu, J.; Gao, N.; Zhu, T.; Wu, J. CFD study of exhaled droplet transmission between occupants under different ventilation strategies in a typical office room. Build. Environ. 2011, 46, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.; Kinzel, M.; Estimating, M. COVID-19 exposure in a classroom setting: A comparison between mathematical and numerical models. Phys. Fluids 2021, 33, 021904. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Galea, E.R.; Grandison, A.; Ewer, J.; Jia, F. A coupled Computational Fluid Dynamics and Wells-Riley model to predict COVID-19 infection probability for passengers on long-distance trains. Saf. Sci. 2022, 147, 105572. [Google Scholar] [CrossRef]
- Su, W.; Yang, B.; Melikov, A.; Liang, C.; Lu, Y.; Wang, F.; Li, A.; Lin, Z.; Li, X.; Cao, G.; et al. Infection probability under different air distribution patterns. Build. Environ. 2022, 207, 108555. [Google Scholar] [CrossRef]
- Riley, E.C.; Murphy, G.; Riley, R.L. Airborne spread of measles in a suburban elementary school. Am. J. Epidemiol. 1978, 107, 421–432. [Google Scholar] [CrossRef]
- Mittal, R.; Meneveau, C.; Wu, W. A mathematical framework for estimating risk of airborne transmission of COVID-19 with application to face mask use and social distancing. Phys. Fluids 2020, 32, 101903. [Google Scholar] [CrossRef]
- Noakes, C.J.; Sleigh, P.A. Applying the Wells-Riley equation to the risk of airborne infection in hospital environments: The importance of stochastic and proximity effects. In Proceedings of the Indoor Air 2008, the 11th International Conference on Indoor Air Quality and Climate, Copenhagen, Denmark, 17–22 August 2008. [Google Scholar]
- Zhang, S.; Lin, Z. Dilution-based evaluation of airborne infection risk-Thorough expansion of Wells-Riley model. Build. Environ. 2021, 194, 107674. [Google Scholar] [CrossRef]
- Shao, X.; Li, X. COVID-19 transmission in the first presidential debate in 2020. Phys. Fluids 2020, 32, 115125. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Sun, Q.; Knopf, J.; Herrmann, M.; Lin, L.; Jiang, J.; Shao, C.; Li, P.; He, X.; et al. Immune response in COVID-19: What is next? Cell Death Differ. 2022, 29, 1107–1122. [Google Scholar] [CrossRef]
- Beck, C.; Cohen, E.G.D. Superstatistics. Phys. A 2003, 322, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Tsallis, C. Possible generalization of Boltzmann-Gibbs statistics. J. Stat. Phys. 1988, 52, 479. [Google Scholar] [CrossRef]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290. [Google Scholar] [CrossRef]
- Gupta, S.L.; Jaiswal, R.K. Neutralizing antibody: A savior in the COVID-19 disease. Mol. Biol. Rep. 2022, 49, 2465–2474. [Google Scholar] [CrossRef]
- Lamers, M.M.; Haagmans, B.L. SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 2022, 20, 270–284. [Google Scholar] [CrossRef]
- Beck, C.; Cohen, E.G.D.; Swinney, H.L. From time series to superstatistics. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2005, 72, 056133. [Google Scholar] [CrossRef] [Green Version]
- Feller, W. An Introduction to Probability Theory and Its Applications; John Wiley: London, UK, 1966; Volume II. [Google Scholar]
- Arnold, B.C. Pareto Distributions; International Cooperative Publishing House: Fairland, MD, USA, 1983. [Google Scholar]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Cheemarla, N.R.; Watkins, T.A.; Mihaylova, V.T.; Wang, B.; Zhao, D.; Wang, G.; Landry, M.L.; Foxman, E.F. Dynamic innate immune response determines susceptibility to SARS-CoV-2 infection and early replication kinetics. J. Exp. Med. 2021, 218, 8. [Google Scholar] [CrossRef]
- Matricardi, P.M.; Dal Negro, R.W.; Nisini, R. The first, holistic immunological model of COVID-19: Implications for prevention, diagnosis, and public health measures. Pediatr. Allergy Immunol. 2020, 5, 454–470. [Google Scholar] [CrossRef]
- Morales-Núñez, J.J.; Muñoz-Valle, J.F.; Torres-Hernández, P.C.; Hernández-Bello, J. Overview of Neutralizing Antibodies and Their Potential in COVID-19. Vaccines 2021, 9, 1376. [Google Scholar] [CrossRef]
- Favresse, J.; Gillot, C.; Di Chiaro, L.; Eucher, C.; Elsen, M.; Van Eeckhoudt, S.; David, C.; Morimont, L.; Dogné, J.M.; Douxfils, J. Neutralizing Antibodies in COVID-19 Patients and Vaccine Recipients after Two Doses of BNT162b2. Viruses 2021, 13, 1364. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.S.; Pedron, I.T. Nonlinear differential equations based on nonextensive Tsallis entropy and physical applications. arXiv 1999, arXiv:Cond.mat/9904023v1. [Google Scholar]
- Brouers, F.; Sotolongo-Costa, O. Generalized fractal kinetics in complex systems (application to biophysics and biotechnology). Phys. A 2006, 368, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Van Rossum, G.; Drake, F.L., Jr. Python Reference Manual; Centrum voor Wiskunde en Informatica: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Dorm, J.R.; Prince, P.J. A family of embedded Runge-Kutta formulae. J. Comput. Appl. Math. 1980, 6, 19–26. [Google Scholar]
- Kim, E.-J.; Tenkès, L.-M.; Hollerbach, R.; Radulescu, O. Far-From-Equilibrium Time Evolution between Two Gamma Distributions. Entropy 2017, 19, 511. [Google Scholar] [CrossRef] [Green Version]
- Adams, W.C. Measurement of Breathing Rate and Volume in Routinely Performed Daily Activities: Final Report, Contract No. A033-205; California Air Resources Board: Sacramento, CA, USA, 1993. [Google Scholar]
- Tsallis, C.; Tirnakli, U. Predicting COVID-19 Peaks Around the World. Front. Phys. 2020, 8, 217. [Google Scholar] [CrossRef]
- Silverman, B.D. Hydrophobic moments of protein structures: Spatially profiling the distribution. Proc. Natl. Acad. Sci. USA 2001, 98, 4996–5001. [Google Scholar] [CrossRef] [Green Version]
- Xenakis, M.N.; Kapetis, D.; Yang, Y.; Heijman, J.; Waxman, S.G.; Lauria, G.; Faber, C.G.; Smeets, H.J.; Westra, R.L.; Lindsey, P.J. Cumulative hydropathic topology of a voltage-gated sodium channel at atomic resolution. Proteins 2020, 88, 1319–1328. [Google Scholar] [CrossRef]
- Xenakis, M.N.; Kapetis, D.; Yang, Y.; Heijman, J.; Waxman, S.G.; Lauria, G.; Faber, C.G.; Smeets, H.J.; Lindsey, P.J.; Westra, R.L. Non-extensitivity and criticality of atomic hydropathicity around a voltage-gated sodium channel’s pore: A modeling study. J. Biol. Phys. 2021, 47, 61–77. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary Param. | Out-of-Host Param. | Within-Host Param. |
|---|---|---|
| (1/h) | N (nr. of occupants) | (VCAPs) |
| (VCAPs/m) | F (nr. of infectors) | (dimensionless) |
| (dimensionless) | V (m) | (1/h) |
| (h) | (m) | |
| (h) | ||
| r (m/h) | ||
| w (VCAPs/h) | ||
| W (m/h) | ||
| (m) | ||
| (h) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xenakis, M.N. Generalizing the Wells–Riley Infection Probability: A Superstatistical Scheme for Indoor Infection Risk Estimation. Entropy 2023, 25, 896. https://doi.org/10.3390/e25060896
Xenakis MN. Generalizing the Wells–Riley Infection Probability: A Superstatistical Scheme for Indoor Infection Risk Estimation. Entropy. 2023; 25(6):896. https://doi.org/10.3390/e25060896
Chicago/Turabian StyleXenakis, Markos N. 2023. "Generalizing the Wells–Riley Infection Probability: A Superstatistical Scheme for Indoor Infection Risk Estimation" Entropy 25, no. 6: 896. https://doi.org/10.3390/e25060896