
Information Encoding in Bursting Spiking Neural Network Modulated by Astrocytes

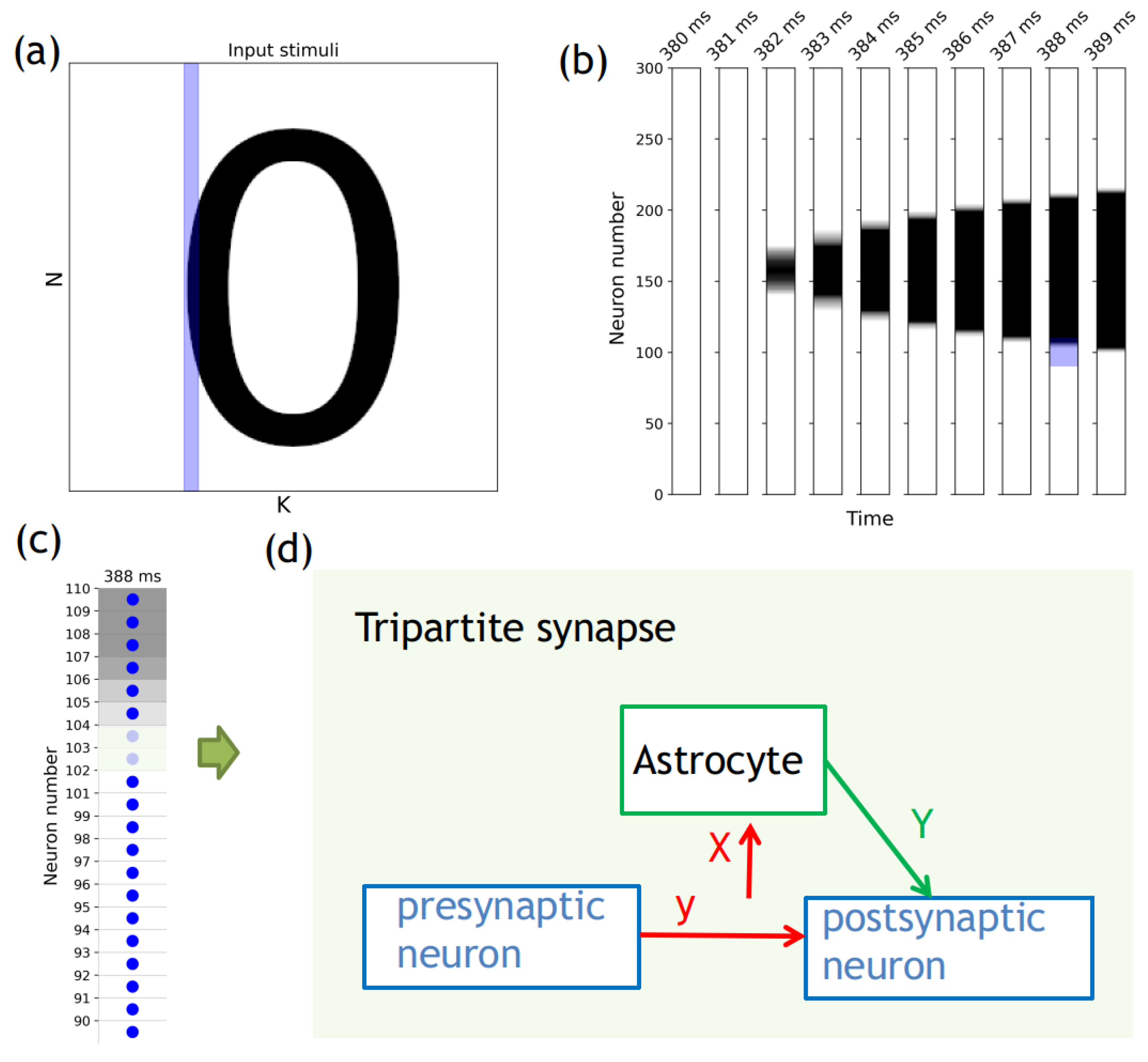{kind=link}
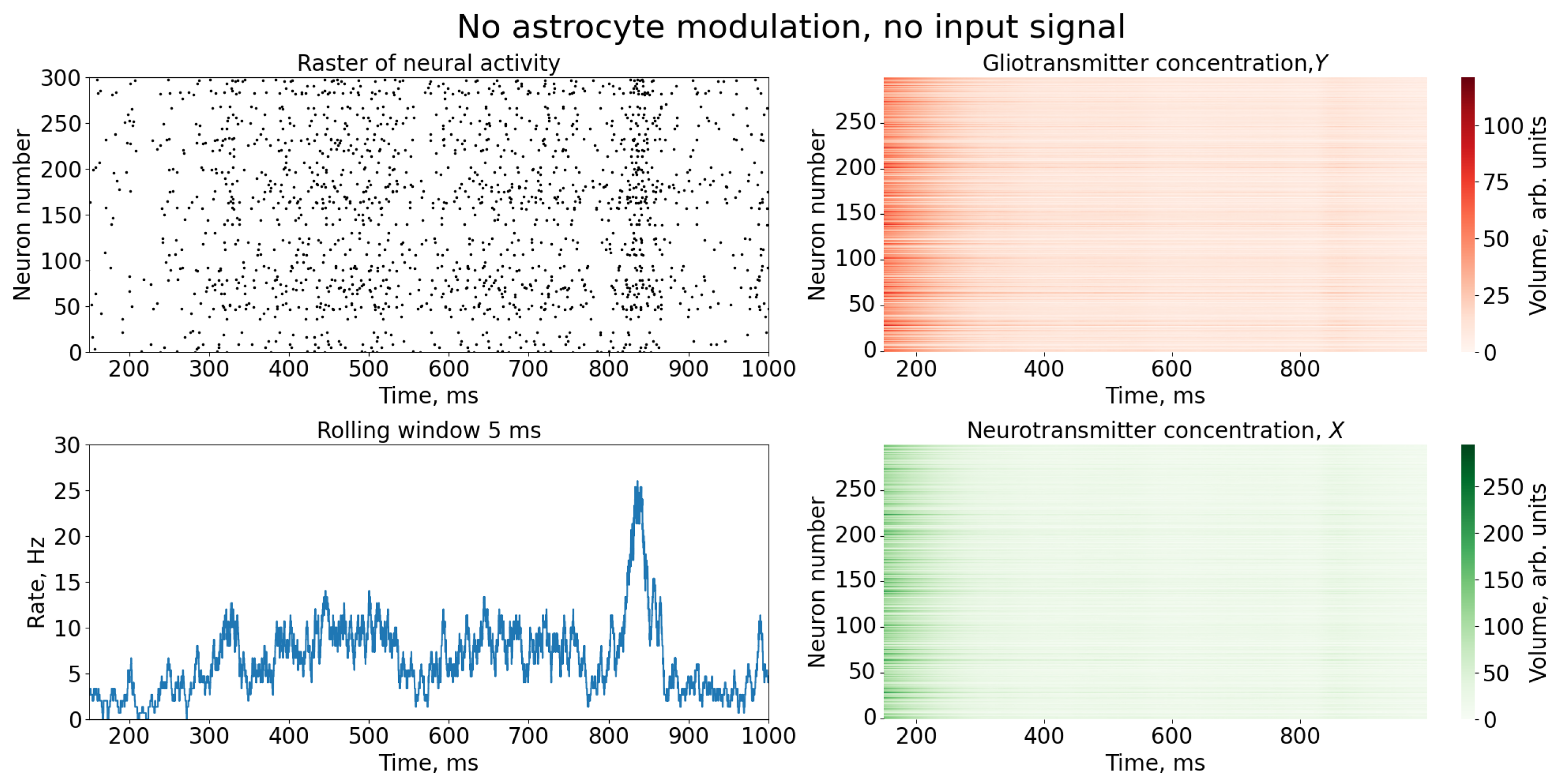{kind=link}
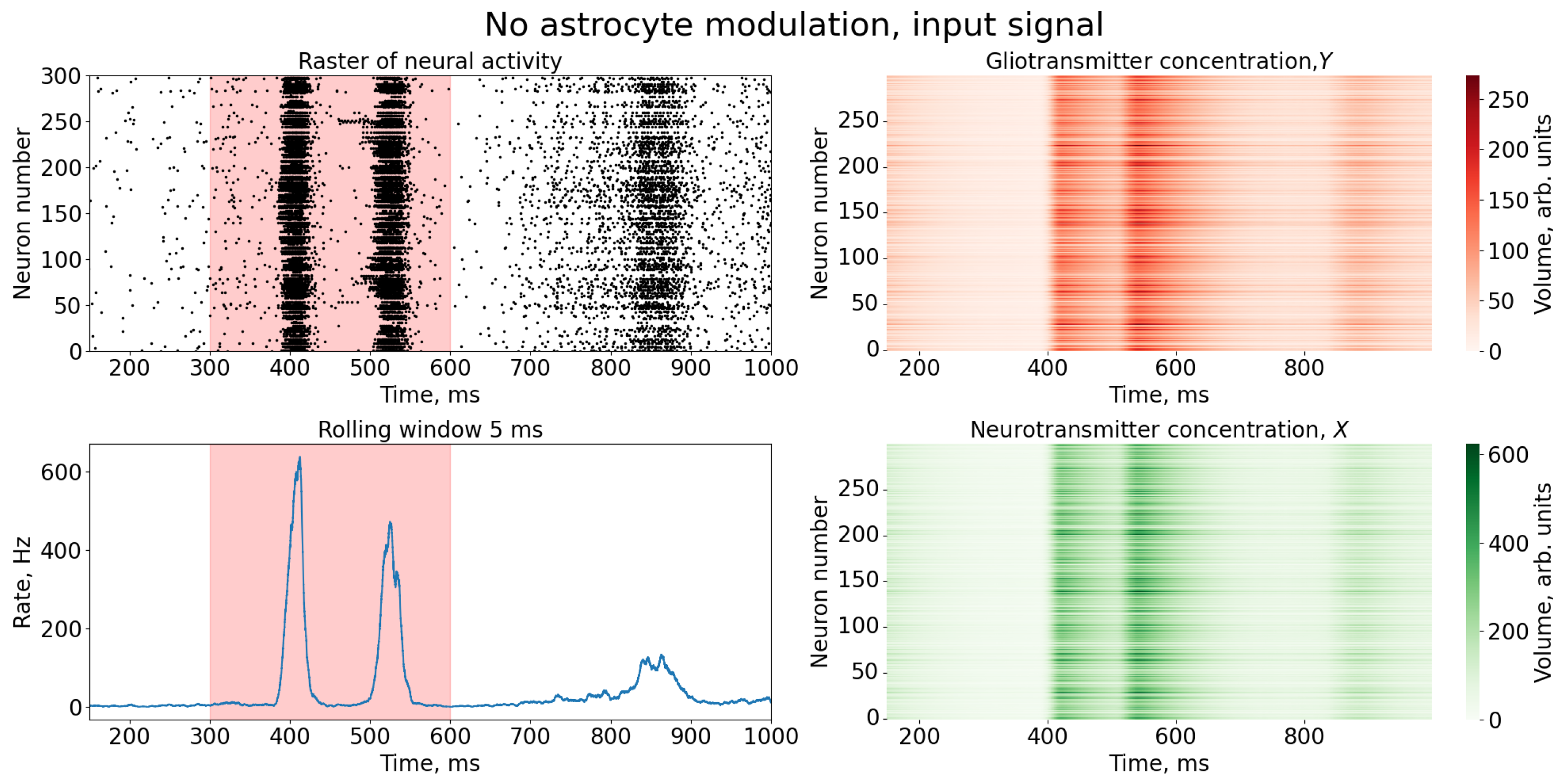{kind=link}
{kind=link}
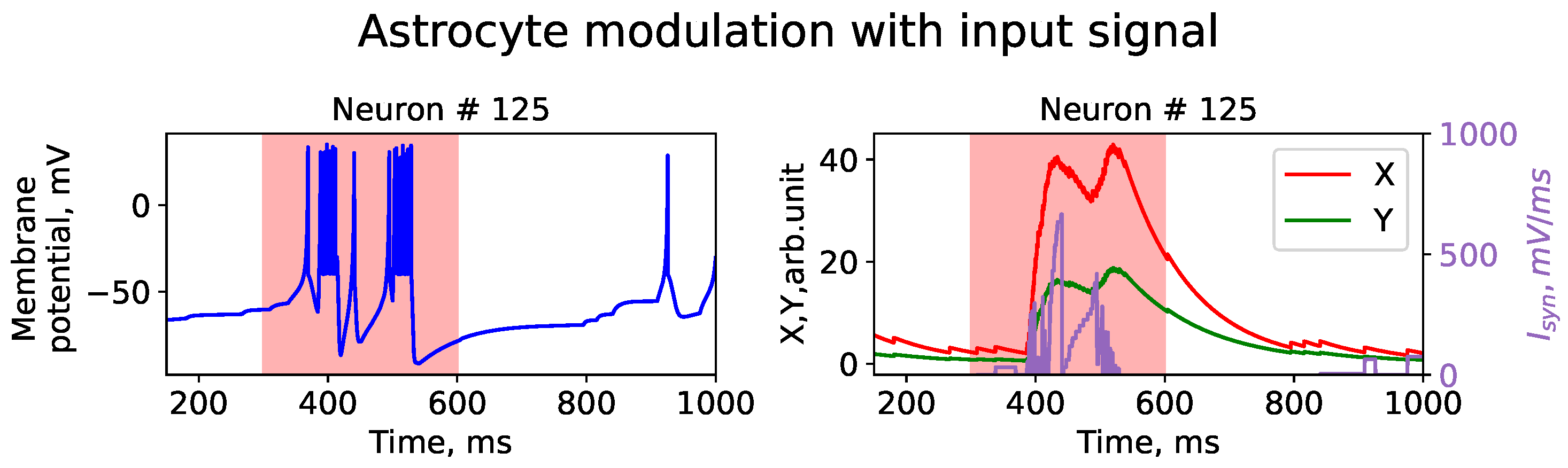{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Model
2.1. Astrocytic Dynamics
2.2. Astrocytic Modulation of Neural Activity
2.3. Stimulation Current
2.4. Neural Network
2.5. Numerical Simulation Method
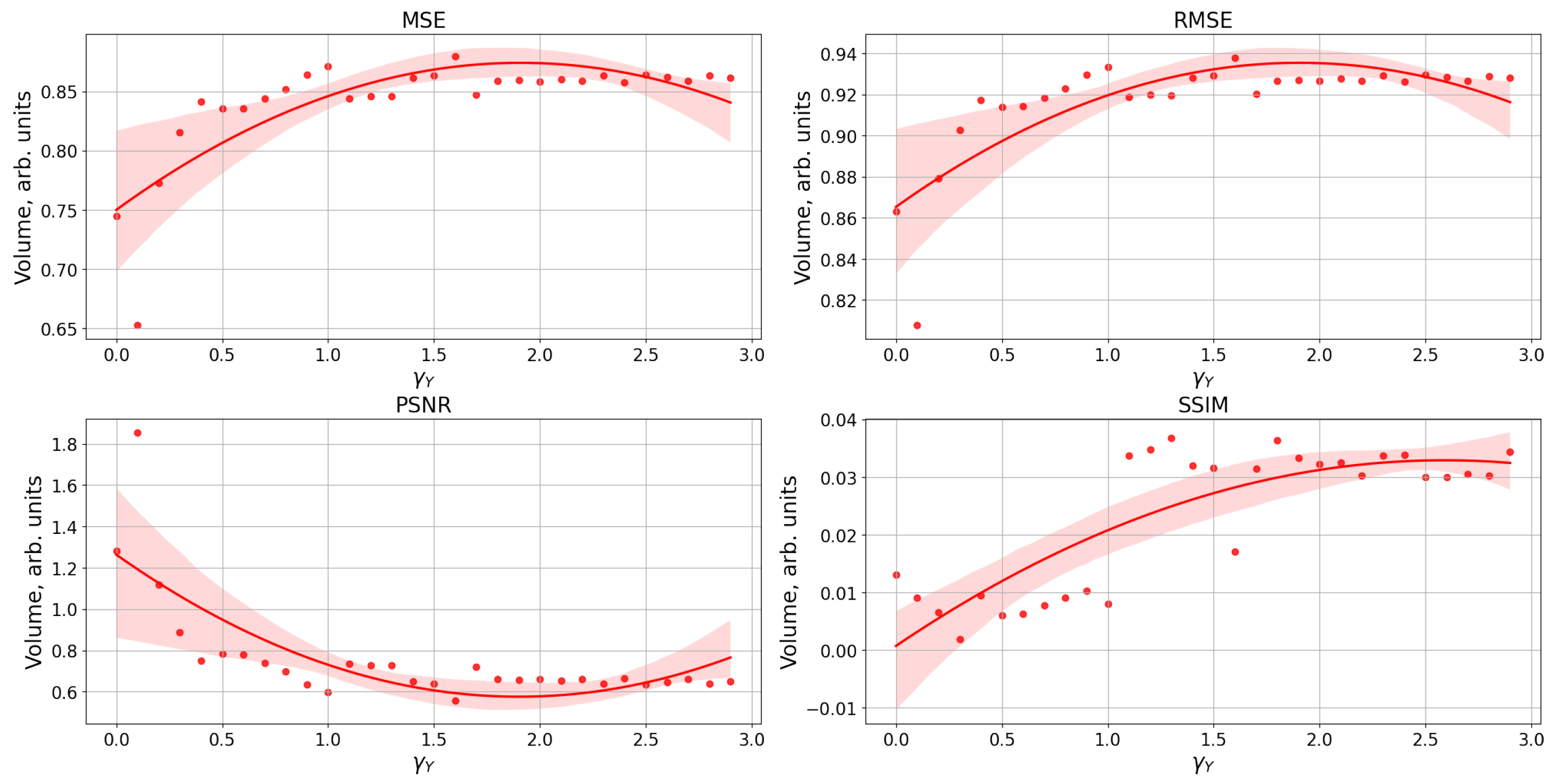
2.6. Image Similarity Metrics
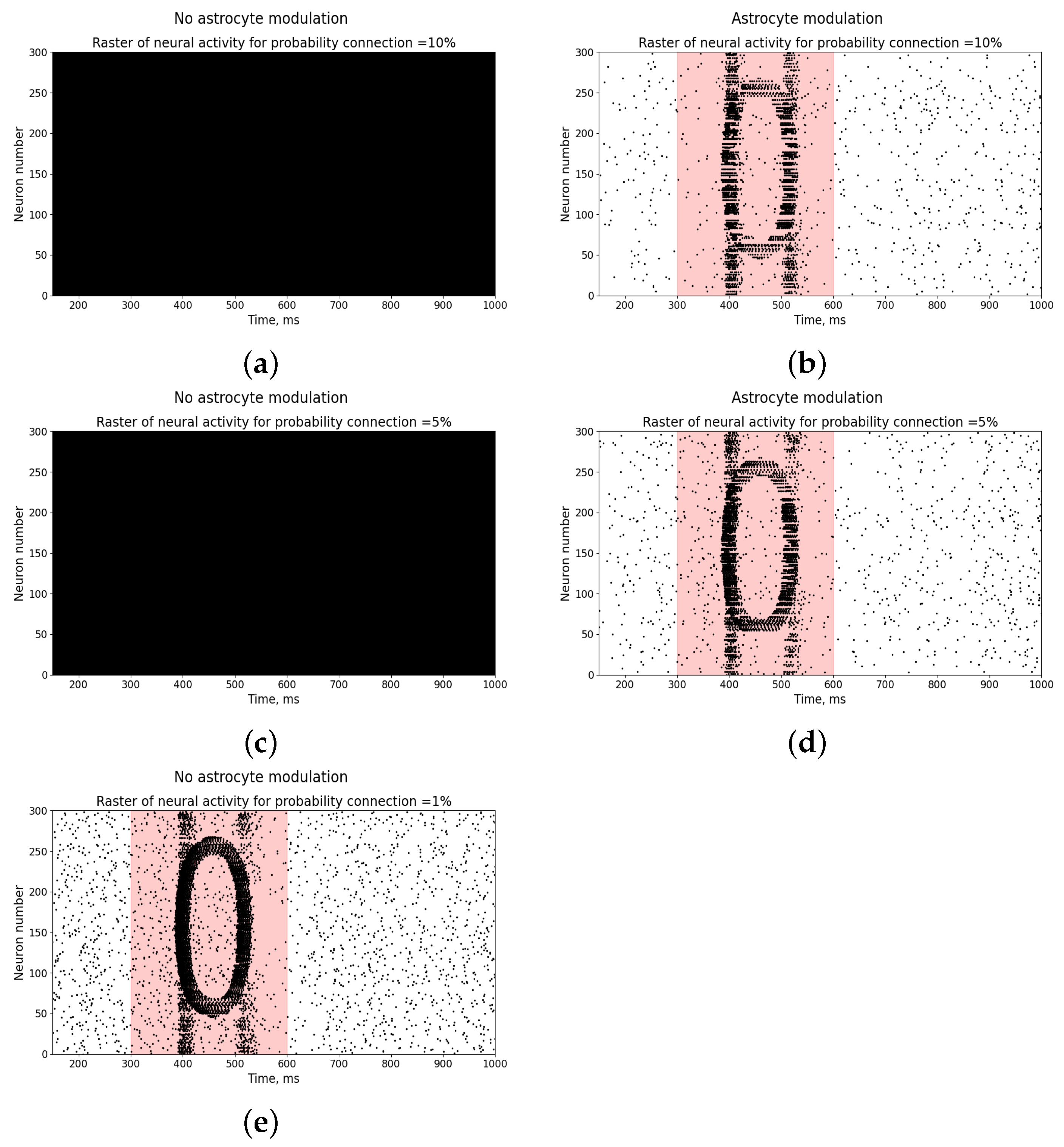
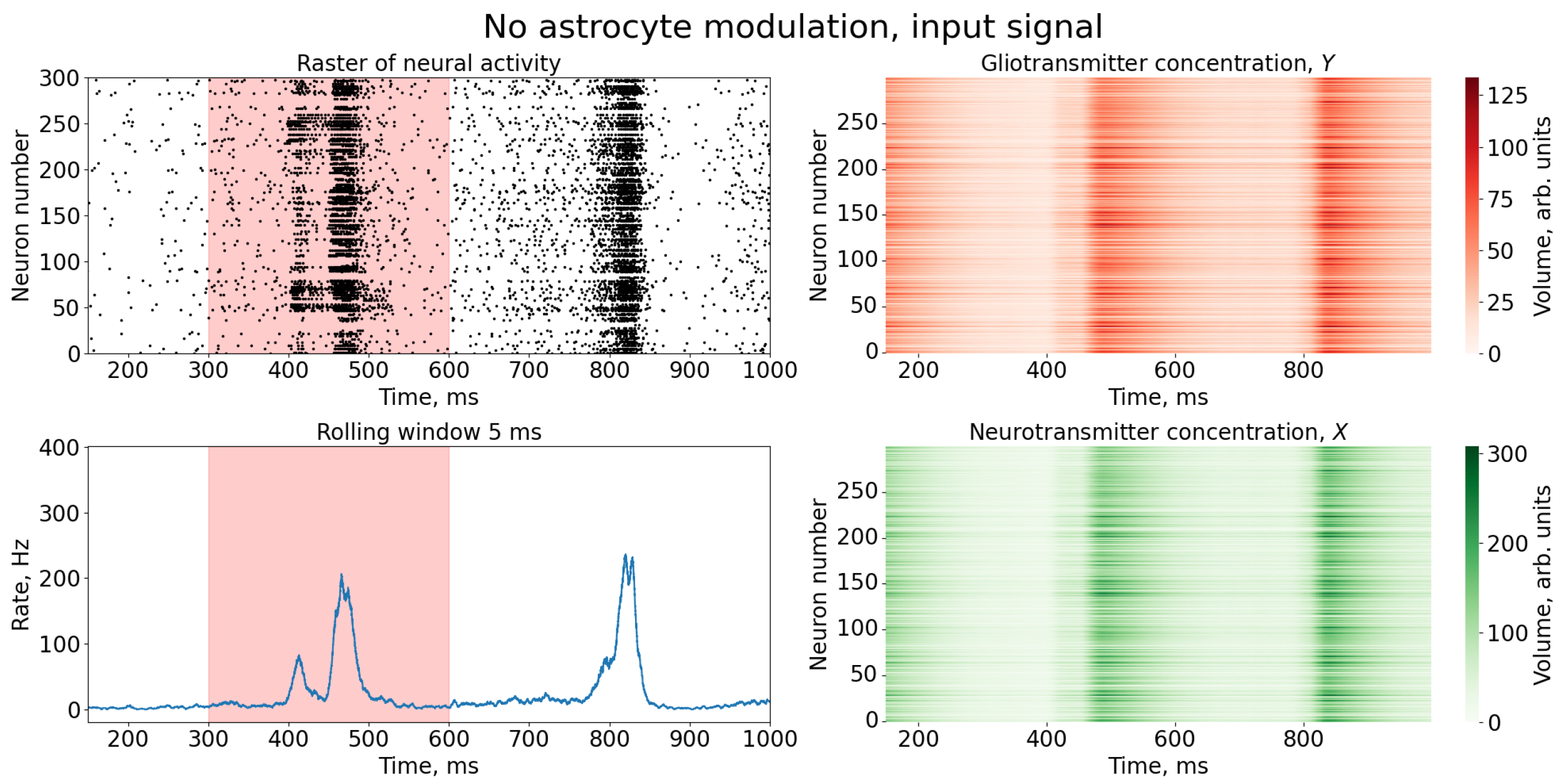
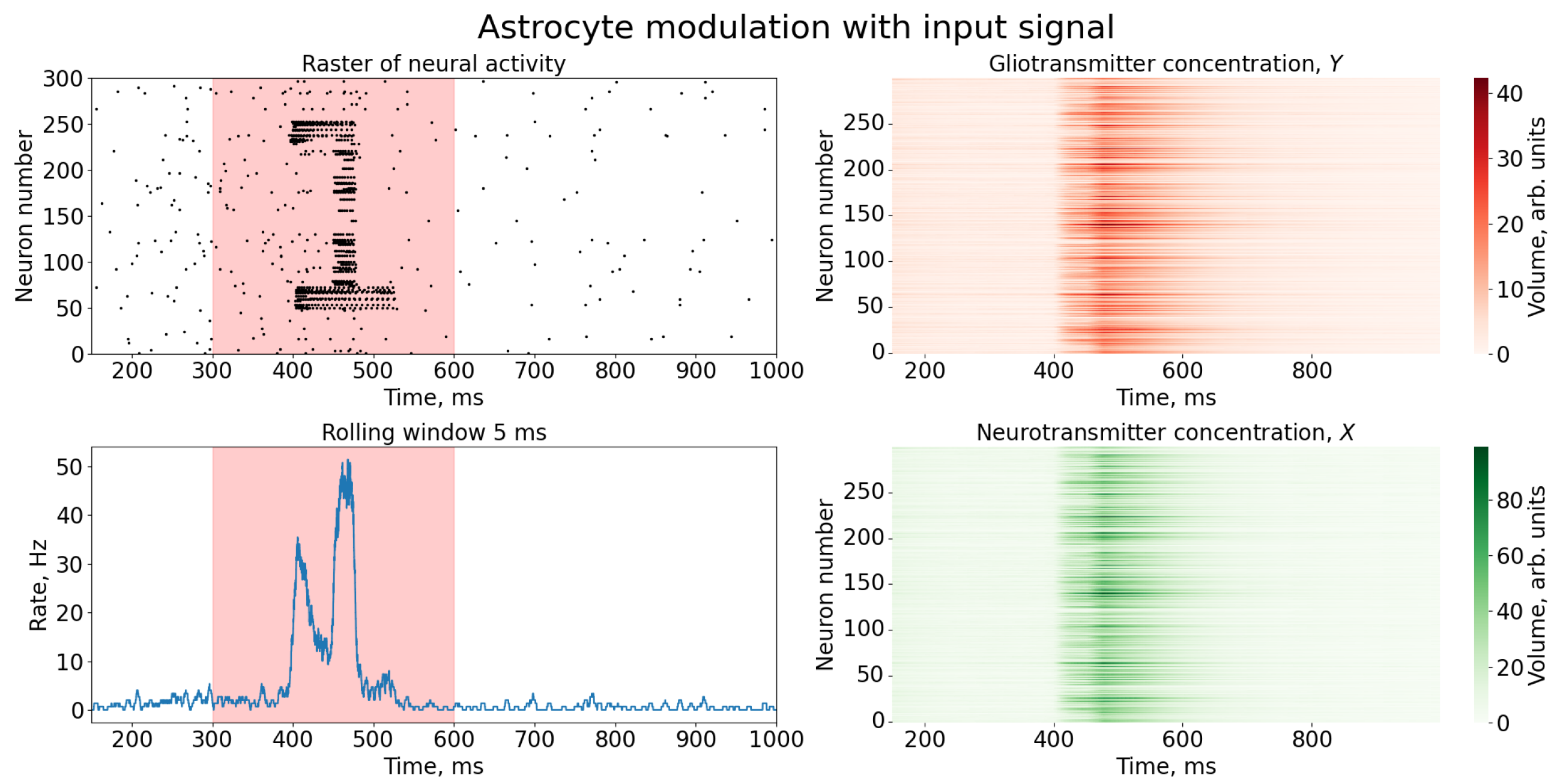
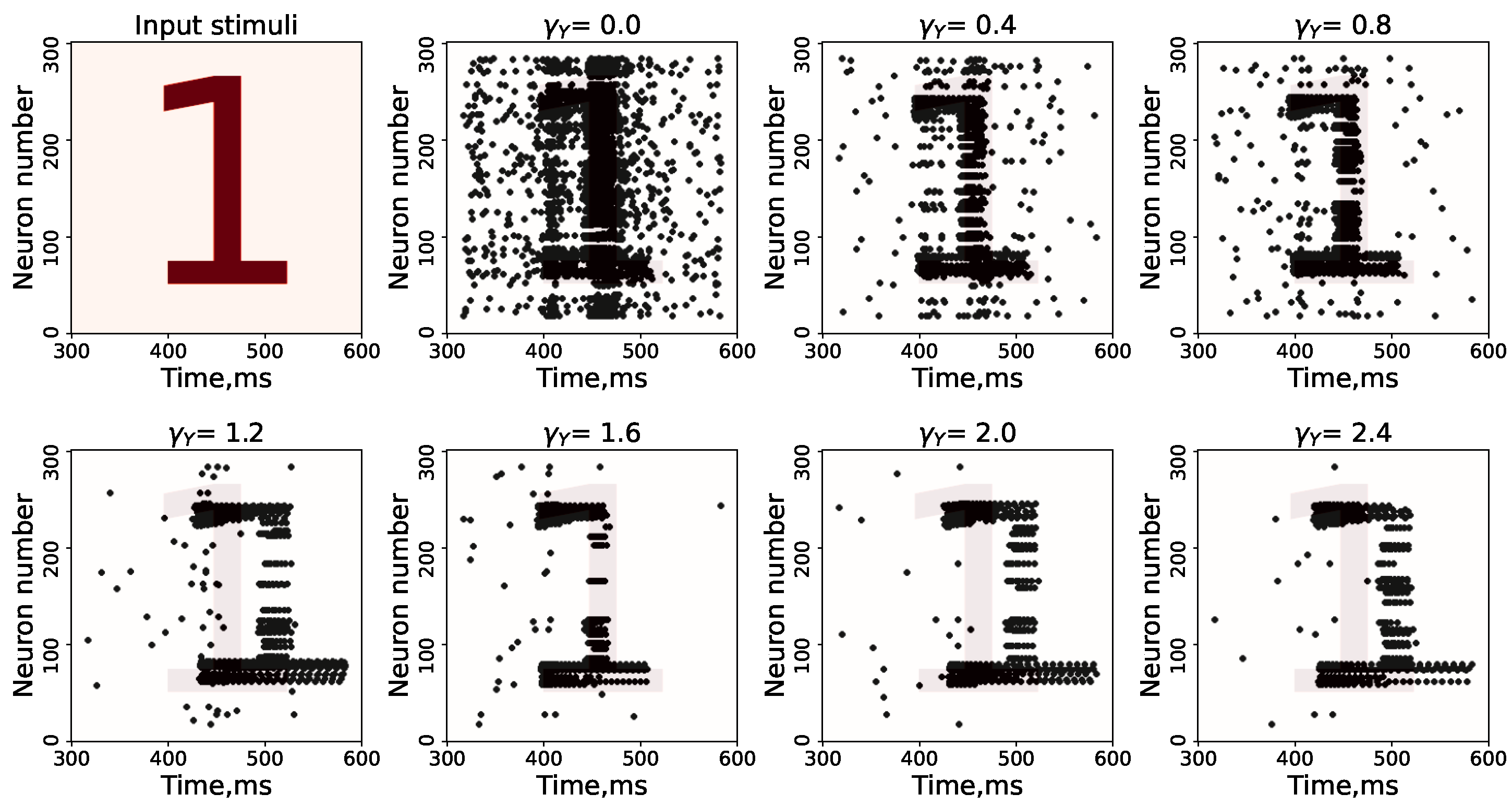
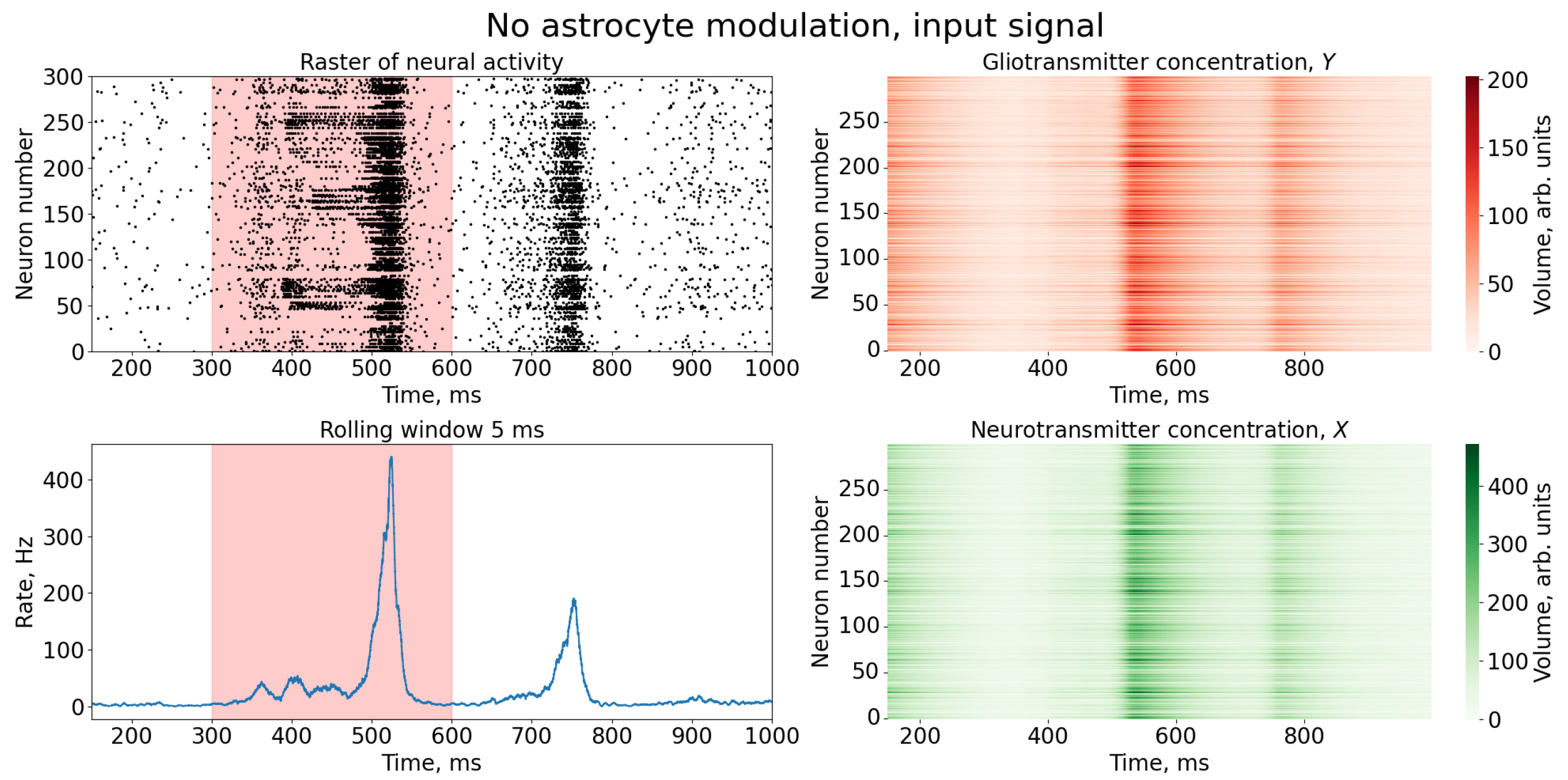
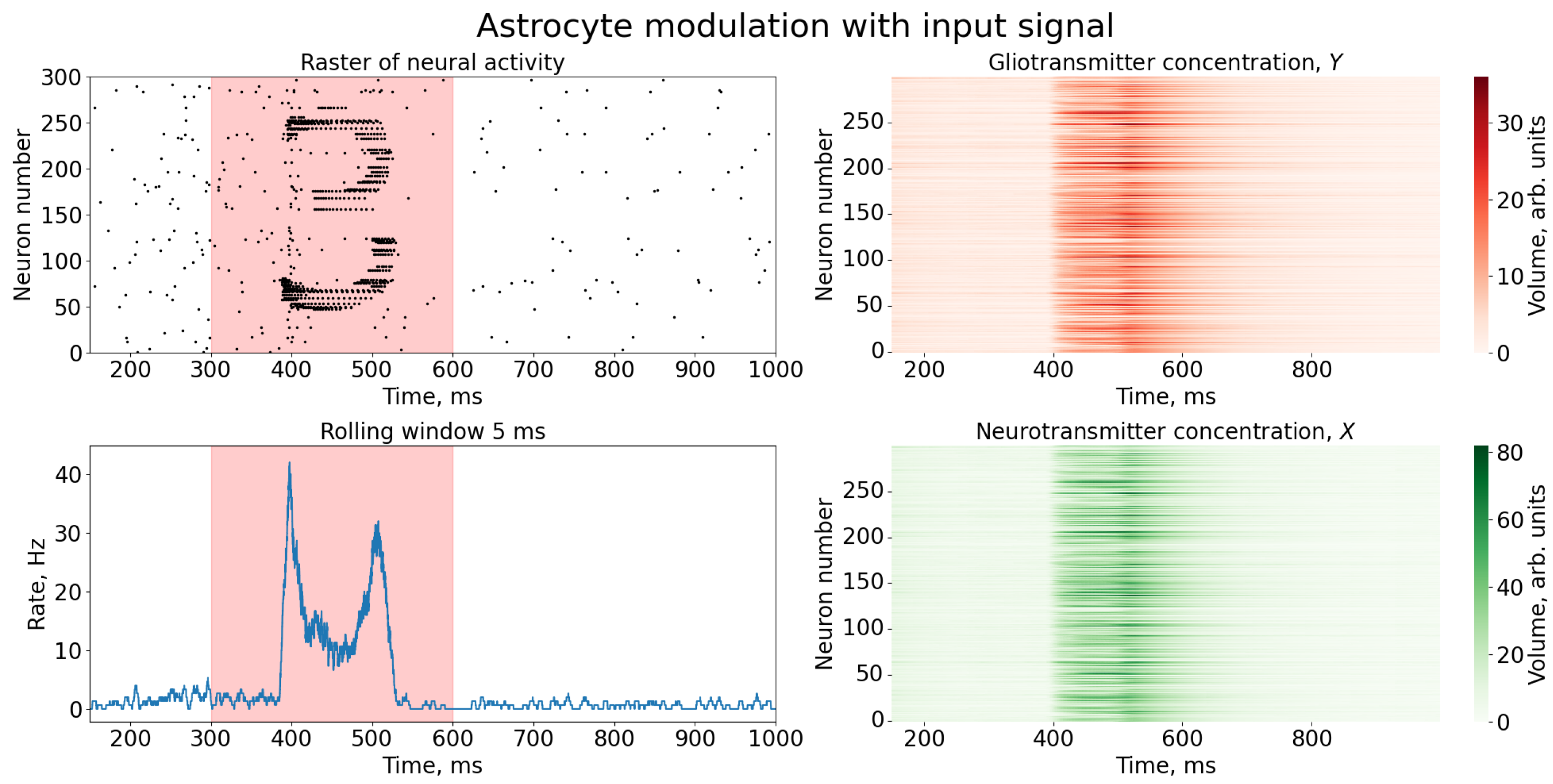
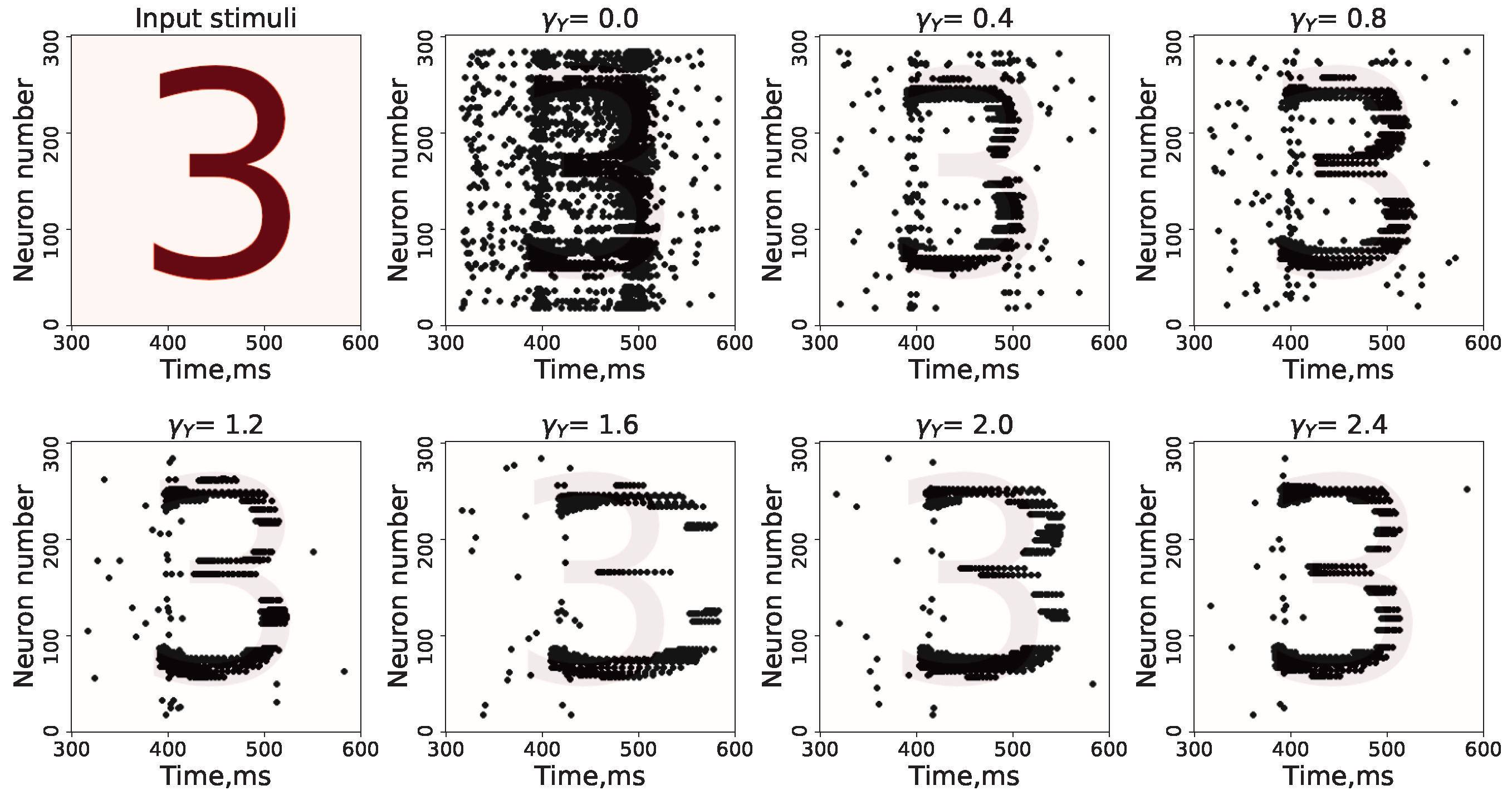
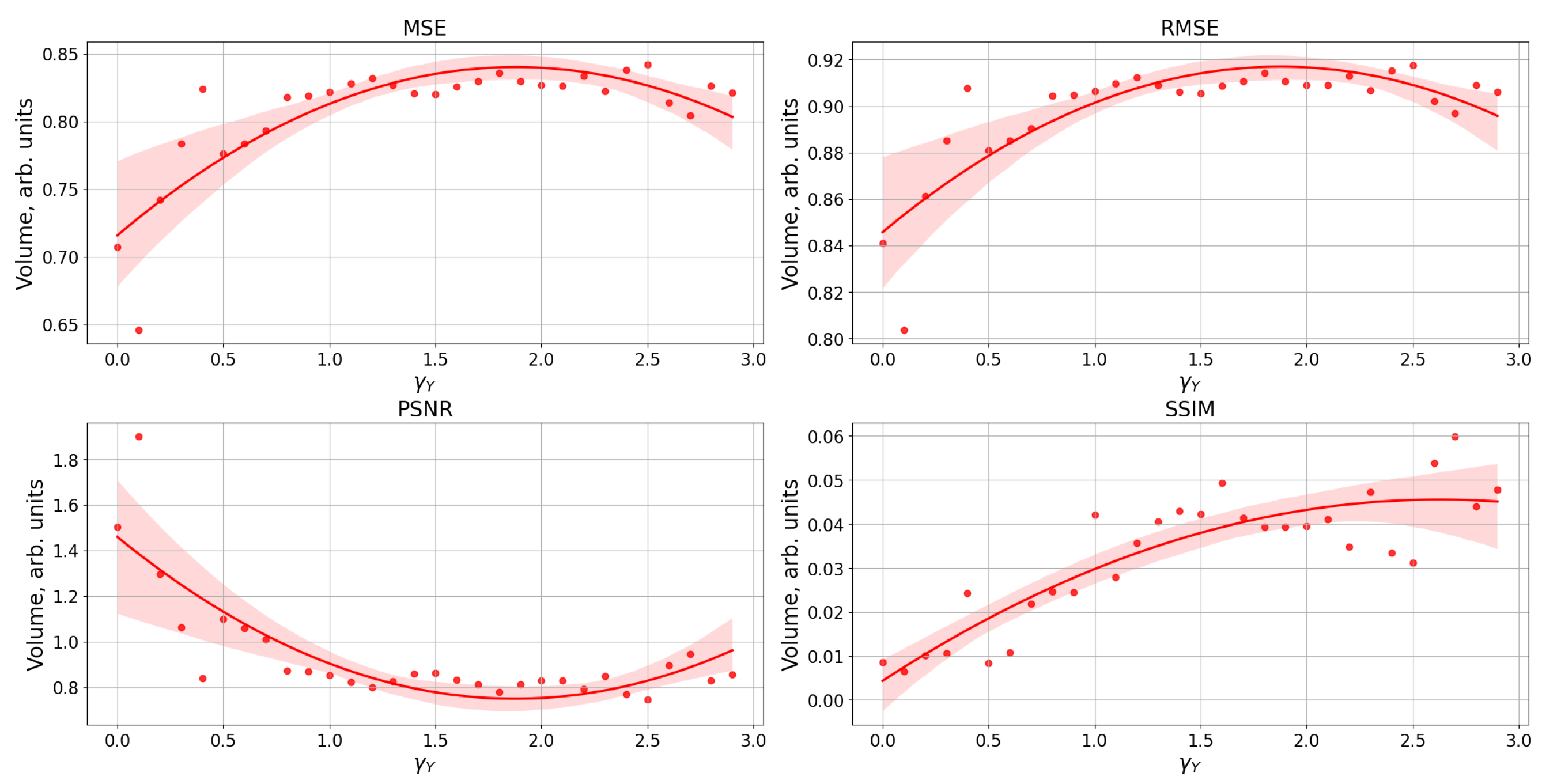
3. Results
Study of Neuron and Neural Network Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Cases with Other Numbers








References
- Ghosh-Dastidar, S.; Adeli, H. Improved spiking neural networks for EEG classification and epilepsy and seizure detection. Integr. Comput.-Aided Eng. 2007, 14, 187–212. [Google Scholar] [CrossRef]
- Dora, S.; Kasabov, N. Spiking Neural Networks for Computational Intelligence: An Overview. Big Data Cogn. Comput. 2021, 5, 67. [Google Scholar] [CrossRef]
- Lobov, S.; Chernyshov, A.; Krilova, N.; Shamshin, M.; Kazantsev, V. Competitive learning in a spiking neural network: Towards an intelligent pattern classifier. Sensors 2020, 20, 500. [Google Scholar] [CrossRef]
- Wagenaar, D.; Pine, J.; Potter, S. An extremely rich repertoire of bursting patterns during the development of cortical cultures. BMC Neurosci. 2006, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kepecs, A.; Lisman, J. Information encoding and computation with spikes and bursts. Netw. Comput. Neural Syst. 2003, 14, 103. [Google Scholar] [CrossRef]
- Kepecs, A.; Wang, X.; Lisman, J. Bursting neurons signal input slope. J. Neurosci. 2002, 22, 9053–9062. [Google Scholar] [CrossRef]
- Prince, D. Neurophysiology of epilepsy. Annu. Rev. Neurosci. 1978, 1, 395–415. [Google Scholar] [CrossRef] [PubMed]
- Pimashkin, A.; Kastalskiy, I.; Simonov, A.; Koryagina, E.; Mukhina, I.; Kazantsev, V. Spiking signatures of spontaneous activity bursts in hippocampal cultures. Front. Comput. Neurosci. 2011, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Feinerman, O.; Segal, M.; Moses, E. Identification and dynamics of spontaneous burst initiation zones in unidimensional neuronal cultures. J. Neurophysiol. 2007, 97, 2937–2948. [Google Scholar] [CrossRef] [PubMed]
- Krahe, R.; Gabbiani, F. Burst firing in sensory systems. Nat. Rev. Neurosci. 2004, 5, 13–23. [Google Scholar] [CrossRef]
- Whitmire, C.; Waiblinger, C.; Schwarz, C.; Stanley, G. Information coding through adaptive gating of synchronized thalamic bursting. Cell Rep. 2016, 14, 795–807. [Google Scholar] [CrossRef]
- Borden, P.; Wright, N.; Morrissette, A.; Jaeger, D.; Haider, B.; Stanley, G. Thalamic bursting and the role of timing and synchrony in thalamocortical signaling in the awake mouse. Neuron 2022, 110, 2836–2853. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Rogawski, M. T-type calcium channels mediate the transition between tonic and phasic firing in thalamic neurons. Proc. Natl. Acad. Sci. USA 1989, 86, 7228–7232. [Google Scholar] [CrossRef]
- Crick, F. Function of the thalamic reticular complex: The searchlight hypothesis. Proc. Natl. Acad. Sci. USA 1984, 81, 4586–4590. [Google Scholar] [CrossRef] [PubMed]
- Lesica, N.; Stanley, G. Encoding of natural scene movies by tonic and burst spikes in the lateral geniculate nucleus. J. Neurosci. 2004, 24, 10731–10740. [Google Scholar] [CrossRef] [PubMed]
- Sherman, S. Dual response modes in lateral geniculate neurons: Mechanisms and functions. Vis. Neurosci. 1996, 13, 205–213. [Google Scholar] [CrossRef]
- Wang, X.; Wei, Y.; Vaingankar, V.; Wang, Q.; Koepsell, K.; Sommer, F.; Hirsch, J. Feedforward excitation and inhibition evoke dual modes of firing in the cat’s visual thalamus during naturalistic viewing. Neuron 2007, 55, 465–478. [Google Scholar] [CrossRef]
- Mukherjee, P.; Kaplan, E. Dynamics of neurons in the cat lateral geniculate nucleus: In vivo electrophysiology and computational modeling. J. Neurophysiol. 1995, 74, 1222–1243. [Google Scholar] [CrossRef]
- Wolfart, J.; Debay, D.; Le Masson, G.; Destexhe, A.; Bal, T. Synaptic background activity controls spike transfer from thalamus to cortex. Nat. Neurosci. 2005, 8, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Sharpee, T.; Sugihara, H.; Kurgansky, A.; Rebrik, S.; Stryker, M.; Miller, K. Adaptive filtering enhances information transmission in visual cortex. Nature 2006, 439, 936–942. [Google Scholar] [CrossRef]
- Wagenaar, D.; Madhavan, R.; Pine, J.; Potter, S. Controlling bursting in cortical cultures with closed-loop multi-electrode stimulation. J. Neurosci. 2005, 25, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.; Christiansen, N.; Sandvig, A.; Sandvig, I. Selective inhibition of excitatory synaptic transmission alters the Emergent Bursting Dyn. Vitr. Neural Networks. Front. Neural Circuits 2023, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Cortes, J.; Desroches, M.; Rodrigues, S.; Veltz, R.; Muñoz, M.; Sejnowski, T. Short-term synaptic plasticity in the deterministic Tsodyks–Markram model leads to unpredictable network dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 16610–16615. [Google Scholar] [CrossRef]
- Fortune, E.; Rose, G. Short-term synaptic plasticity as a temporal filter. Trends Neurosci. 2001, 24, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Tsodyks, M.; Wu, S. Short-term synaptic plasticity. Scholarpedia 2013, 8, 3153. [Google Scholar] [CrossRef]
- Markram, H.; Muller, E.; Ramaswamy, S.; Reimann, M.; Abdellah, M.; Sanchez, C.; Ailamaki, A.; Alonso-Nanclares, L.; Antille, N.; Arsever, S. Others Reconstruction and simulation of neocortical microcircuitry. Cell 2015, 163, 456–492. [Google Scholar] [CrossRef]
- Billeh, Y.; Cai, B.; Gratiy, S.; Dai, K.; Iyer, R.; Gouwens, N.; Abbasi-Asl, R.; Jia, X.; Siegle, J.; Olsen, S. Others Systematic integration of structural and functional data into multi-scale models of mouse primary visual cortex. Neuron 2020, 106, 388–403. [Google Scholar] [CrossRef]
- Antolik, J.; Hofer, S.; Bednar, J.; Mrsic-Flogel, T. Model constrained by visual hierarchy improves prediction of neural responses to natural scenes. PLoS Comput. Biol. 2016, 12, e1004927. [Google Scholar] [CrossRef]
- Chizhov, A. Conductance-based refractory density model of primary visual cortex. J. Comput. Neurosci. 2014, 36, 297–319. [Google Scholar] [CrossRef]
- Potjans, T.; Diesmann, M. The cell-type specific cortical microcircuit: Relating structure and activity in a full-scale spiking network model. Cereb. Cortex 2014, 24, 785–806. [Google Scholar] [CrossRef]
- Kazantsev, V.; Gordleeva, S.; Stasenko, S.; Dityatev, A. A Homeostatic Model of Neuronal Firing Governed by Feedback Signals from the Extracellular Matrix; Public Library of Science: San Francisco, CA, USA, 2012. [Google Scholar]
- Lazarevich, I.; Stasenko, S.; Rozhnova, M.; Pankratova, E.; Dityatev, A.; Kazantsev, V. Activity-dependent switches between dynamic regimes of extracellular matrix expression. PLoS ONE 2020, 15, e0227917. [Google Scholar] [CrossRef] [PubMed]
- Rozhnova, M.; Pankratova, E.; Stasenko, S.; Kazantsev, V. Bifurcation analysis of multistability and oscillation emergence in a model of brain extracellular matrix. Chaos Solitons Fractals 2021, 151, 111253. [Google Scholar] [CrossRef]
- Stasenko, S.; Kazantsev, V. Bursting Dynamics of Spiking Neural Network Induced by Active Extracellular Medium. Mathematics 2023, 11, 2109. [Google Scholar] [CrossRef]
- Oschmann, F.; Berry, H.; Obermayer, K.; Lenk, K. From in silico astrocyte cell models to neuron-astrocyte network models: A review. Brain Res. Bull. 2018, 136, 76–84. [Google Scholar] [CrossRef]
- Halassa, M.; Haydon, P. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Annu. Rev. Physiol. 2010, 72, 335–355. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Araque, A. Astrocyte regulation of neural circuit activity and network states. Glia 2022, 70, 1455–1466. [Google Scholar] [CrossRef]
- Gordleeva, S.; Tsybina, Y.; Krivonosov, M.; Ivanchenko, M.; Zaikin, A.; Kazantsev, V.; Gorban, A. Modeling working memory in a spiking neuron network accompanied by astrocytes. Front. Cell. Neurosci. 2021, 15, 631485. [Google Scholar] [CrossRef]
- Abrego, L.; Gordleeva, S.; Kanakov, O.; Krivonosov, M.; Zaikin, A. Estimating integrated information in bidirectional neuron-astrocyte communication. Phys. Rev. E 2021, 103, 022410. [Google Scholar] [CrossRef]
- Tsybina, Y.; Kastalskiy, I.; Krivonosov, M.; Zaikin, A.; Kazantsev, V.; Gorban, A.; Gordleeva, S. Astrocytes mediate analogous memory in a multi-layer neuron–astrocyte network. Neural Comput. Appl. 2022, 34, 9147–9160. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.; Haydon, P. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur. J. Neurosci. 1998, 10, 2129–2142. Available online: http://www.ncbi.nlm.nih.gov/pubmed/9753099 (accessed on 20 February 2023). [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.; Haydon, P. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999, 22, 208–215. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0166223698013496 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Wittenberg, G.; Sullivan, M.; Tsien, J. Synaptic reentry reinforcement based network model for long-term memory consolidation. Hippocampus 2002, 12, 637–647. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12440578 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Wang, X. Synaptic basis of cortical persistent activity: The importance of NMDA receptors to working memory. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 9587–9603. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10531461 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Haydon, P. GLIA: Listening and talking to the synapse. Nat. Rev. Neurosci. 2001, 2, 185–193. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11256079 (accessed on 20 February 2023). [CrossRef]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Martín, E.; Fernández, M.; Perea, G.; Pascual, O.; Haydon, P.; Araque, A.; Ceña, V. Adenosine released by astrocytes contributes to hypoxia-induced modulation of synaptic transmission. Glia 2007, 55, 36–45. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17004232 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Kumar, R.; Huang, Y.; Chen, C.; Tzeng, S.; Chan, C. Astrocytic regulation of synchronous bursting in cortical cultures: From local to global. Cereb. Cortex Commun. 2020, 1, tgaa053. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.; Wall, T.; Sims, R.; Neale, S.; Nisenbaum, E.; Parri, H.; Salt, T. Astrocytes modulate thalamic sensory processing via mGlu2 receptor activation. Neuropharmacology 2017, 121, 100–110. [Google Scholar] [CrossRef]
- Kwak, H.; Koh, W.; Kim, S.; Song, K.; Shin, J.; Lee, J.; Lee, E.; Bae, J.; Ha, G.; Oh, J. Others Astrocytes control sensory acuity via tonic inhibition in the thalamus. Neuron 2020, 108, 691–706. [Google Scholar] [CrossRef]
- Nadkarni, S.; Jung, P. Dressed neurons: Modeling neural-glial interactions. Phys. Biol. 2004, 1, 35–41. Available online: http://www.ncbi.nlm.nih.gov/pubmed/16204820 (accessed on 20 February 2023). [CrossRef]
- Nadkarni, S.; Jung, P. Modeling synaptic transmission of the tripartite synapse. Phys. Biol. 2007, 4, 1–9. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17406080 (accessed on 20 February 2023). [CrossRef]
- Volman, V.; Ben-Jacob, E.; Levine, H. The astrocyte as a gatekeeper of synaptic information transfer. Neural Comput. 2007, 326, 303–326. Available online: http://www.mitpressjournals.org/doi/abs/10.1162/neco.2007.19.2.303 (accessed on 20 February 2023). [CrossRef] [PubMed]
- De Pitta, M.; Volman, V.; Berry, H.; Ben-Jacob, E. A tale of two stories: Astrocyte regulation of synaptic depression and facilitation. PloS Comput. Biol. 2011, 7, e1002293. Available online: http://dx.plos.org/10.1371/journal.pcbi.1002293 (accessed on 20 February 2023). [CrossRef]
- Postnov, D.; Ryazanova, L.; Sosnovtseva, O. Functional modeling of neural-glial interaction. Bio-System 2007, 89, 84–91. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17320272 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Amiri, M.; Bahrami, F.; Janahmadi, M. Functional contributions of astrocytes in synchronization of a neuronal network model. J. Theor. Biol. 2009, 292C, 60–70. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21978738 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Wade, J.; McDaid, L.; Harkin, J.; Crunelli, V.; Kelso, J. Bidirectional Coupling between Astrocytes and Neurons Mediates Learning and Dynamic Coordination in the Brain: A Multiple Modeling Approach. PLoS ONE 2011, 6, e29445. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Hosseinmardi, N.; Bahrami, F.; Janahmadi, M. Astrocyte-neuron interaction as a mechanism responsible for generation of neural synchrony: A study based on modeling and experiments. J. Comput. Neurosci. 2013, 34, 489–504. [Google Scholar] [CrossRef]
- Pankratova, E.; Kalyakulina, A.; Stasenko, S.; Gordleeva, S.; Lazarevich, I.; Kazantsev, V. Neuronal synchronization enhanced by neuron–astrocyte interaction. Nonlinear Dyn. 2019, 97, 647–662. [Google Scholar] [CrossRef]
- Stasenko, S.; Hramov, A.; Kazantsev, V. Loss of neuron network coherence induced by virus-infected astrocytes: A model study. Sci. Rep. 2023, 13, 6401. [Google Scholar] [CrossRef]
- Stasenko, S.; Kazantsev, V. Dynamic Image Representation in a Spiking Neural Network Supplied by Astrocytes. Mathematics 2023, 11, 561. [Google Scholar] [CrossRef]
- Stasenko, S.; Kazantsev, V. Astrocytes Enhance Image Representation Encoded in Spiking Neural Network. In Proceedings of the Advances In Neural Computation, Machine Learning, And Cognitive Research VI: Selected Papers From The XXIV International Conference On Neuroinformatics, Moscow, Russia, 17–21 October 2022; pp. 200–206. [Google Scholar]
- Gordleeva, S.; Stasenko, S.; Semyanov, A.; Dityatev, A.; Kazantsev, V. Bi-directional astrocytic regulation of neuronal activity within a network. Front. Comput. Neurosci. 2012, 6, 92. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3487184/ (accessed on 20 February 2023). [CrossRef]
- De Pittà, M. Gliotransmitter exocytosis and its consequences on synaptic transmission. Comput. Gliosci. 2019, 245–287. [Google Scholar] [CrossRef]
- Lenk, K.; Satuvuori, E.; Lallouette, J.; Guevara, A.; Berry, H.; Hyttinen, J. A computational model of interactions between neuronal and astrocytic networks: The role of astrocytes in the stability of the neuronal firing rate. Front. Comput. Neurosci. 2020, 13, 92. [Google Scholar] [CrossRef]
- Lazarevich, I.; Stasenko, S.; Kazantsev, V. Synaptic multistability and network synchronization induced by the neuron–glial interaction in the brain. JETP Lett. 2017, 105, 210–213. [Google Scholar] [CrossRef]
- Stasenko, S.; Lazarevich, I.; Kazantsev, V. Quasi-synchronous neuronal activity of the network induced by astrocytes. Procedia Comput. Sci. 2020, 169, 704–709. [Google Scholar] [CrossRef]
- Barabash, N.; Levanova, T.; Stasenko, S. STSP model with neuron-glial interaction produced bursting activity. In Proceedings of the 2021 Third International Conference Neurotechnologies And Neurointerfaces (CNN), Kaliningrad, Russia, 13–15 September 2021; pp. 12–15. [Google Scholar]
- Stasenko, S.; Kazantsev, V. 3D model of bursting activity generation. In Proceedings of the 2022 Fourth International Conference Neurotechnologies and Neurointerfaces (CNN), Kaliningrad, Russia, 14–16 September 2022; pp. 176–179. [Google Scholar]
- Barabash, N.; Levanova, T.; Stasenko, S. Rhythmogenesis in the mean field model of the neuron–glial network. Eur. Phys. J. Spec. Top. 2023, 1–6. [Google Scholar] [CrossRef]
- Postnov, D.; Koreshkov, R.; Brazhe, N.; Brazhe, A.; Sosnovtseva, O. Dynamical patterns of calcium signaling in a functional model of neuron–astrocyte networks. J. Biol. Phys. 2009, 35, 425–445. [Google Scholar] [CrossRef] [PubMed]
- De Pittà, M.; Brunel, N. Multiple forms of working memory emerge from synapse–astrocyte interactions in a neuron–glia network model. Proc. Natl. Acad. Sci. USA 2022, 119, e2207912119. [Google Scholar] [CrossRef]
- Blum Moyse, L.; Berry, H. Modelling the modulation of cortical Up-Down state switching by astrocytes. PLoS Comput. Biol. 2022, 18, e1010296. [Google Scholar] [CrossRef]
- Izhikevich, E. Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting; The MIT Press: Cambridge, MA, USA, 2007; Volume 441. [Google Scholar] [CrossRef]
- Postnov, D.; Zhirin, R.; Serdobintseva, Y. Noise-induced coherent firing patterns in small neural ensembles with ionic coupling. Izv. VUZ. Appl. Nonlinear Dyn. 2008, 16, 83–100. [Google Scholar]
- Izhikevich, E. Which model to use for cortical spiking neurons? IEEE Trans. Neural Netw. 2004, 15, 1063–1070. [Google Scholar] [CrossRef]
- Angulo, M.; Kozlov, A. Glutamate released from glial cells synchronizes neuronal activity in the hippocampus. J. Neurosci. 2004, 24, 6920–6927. Available online: http://www.jneurosci.org/content/24/31/6920.short (accessed on 20 February 2023). [CrossRef]
- Halassa, M.; Fellin, T.; Haydon, P. Tripartite synapses: Roles for astrocytic purines in the control of synaptic physiology and behavior. Neuropharmacology 2009, 57, 343–346. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19577581 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Perea, G.; Araque, A. Astrocytes potentiate transmitter release at single hippocampal synapses. Science 2007, 317, 1083–1086. Available online: http://www.ncbi.nlm.nih.gov/pubmed/17717185 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Jourdain, P.; Bergersen, L.; Bhaukaurally, K.; Bezzi, P.; Santello, M.; Domercq, M.; Matute, C.; Tonello, F.; Gundersen, V.; Volterra, A. Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 2007, 10, 331–339. Available online: http://www.nature.com/articles/nn1849 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Fiacco, T.; McCarthy, K. Intracellular astrocyte calcium waves in situ increase the frequency of spontaneous AMPA receptor currents in CA1 pyramidal neurons. J. Neurosci. 2004, 24, 722–732. Available online: http://www.jneurosci.org/content/24/3/722.short (accessed on 20 February 2023). [CrossRef]
- Lecarme, O.; Delvare, K. The Book of GIMP: A Complete Guide to Nearly Everything; No Starch Press: San Francisco, CA, USA, 2013. [Google Scholar]
- Winer, J.; Larue, D. Populations of GABAergic neurons and axons in layer I of rat auditory cortex. Neuroscience 1989, 33, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, L.; Villers-Sidani, E. Trajectory of the main GABAergic interneuron populations from early development to old age in the rat primary auditory cortex. Front. Neuroanat. 2014, 8, 40. [Google Scholar] [CrossRef]
- Braitenberg, V.; Schüz, A. Cortex: Statistics and Geometry of Neuronal Connectivity; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Binzegger, T.; Douglas, R.; Martin, K. A quantitative map of the circuit of cat primary visual cortex. J. Neurosci. 2004, 24, 8441–8453. [Google Scholar] [CrossRef]
- Collins, C.; Airey, D.; Young, N.; Leitch, D.; Kaas, J. Neuron densities vary across and within cortical areas in primates. Proc. Natl. Acad. Sci. USA 2010, 107, 15927–15932. [Google Scholar] [CrossRef]
- Alreja, A.; Nemenman, I.; Rozell, C. Constrained brain volume in an efficient coding model explains the fraction of excitatory and inhibitory neurons in sensory cortices. PLoS Comput. Biol. 2022, 18, e1009642. [Google Scholar] [CrossRef] [PubMed]
- Stimberg, M.; Goodman, D.; Brette, R.; Pittà, M. Modeling neuron–glia interactions with the Brian 2 simulator. In Computational Glioscience; Springer: Cham, Switzerland, 2019; pp. 471–505. [Google Scholar] [CrossRef]
- Rusakov, D.; Kullmann, D. Extrasynaptic glutamate diffusion in the hippocampus: Ultrastructural constraints, uptake, and receptor activation. J. Neurosci. 1998, 18, 3158–3170. Available online: https://www.jneurosci.org/lookup/doi/10.1523/JNEUROSCI.18-09-03158.1998 (accessed on 20 February 2023). [CrossRef] [PubMed]
- Van Rossum, G.; Drake, F., Jr. Python Tutorial; Centrum voor Wiskunde en Informatica: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Stimberg, M.; Brette, R.; Goodman, D. Brian 2, an intuitive and efficient neural simulator. eLife 2019, 8, e47314. [Google Scholar] [CrossRef]
- Bisong, E. Matplotlib and seaborn. In Building Machine Learning and Deep Learning Models on Google Cloud Platform: A Comprehensive Guide for Beginners; Apress: Berkeley, CA, USA, 2019; pp. 151–165. [Google Scholar]
- Sara, U.; Akter, M.; Uddin, M. Image quality assessment through FSIM, SSIM, MSE and PSNR—A comparative study. J. Comput. Commun. 2019, 7, 8–18. [Google Scholar] [CrossRef]
- Lu, Y. The level weighted structural similarity loss: A step away from MSE. Proc. AAAI Conf. Artif. Intell. 2019, 33, 9989–9990. [Google Scholar] [CrossRef]
- Søgaard, J.; Krasula, L.; Shahid, M.; Temel, D.; Brunnström, K.; Razaak, M. Applicability of existing objective metrics of perceptual quality for adaptive video streaming. In Electronic Imaging, Image Quality And System Performance XIII; Society for Imaging Science and Technology: Springfield, VA, USA, 2016. [Google Scholar]
- Deshpande, R.; Ragha, L.; Sharma, S. Video quality assessment through PSNR estimation for different compression standards. Indones. J. Electr. Eng. Comput. Sci. 2018, 11, 918–924. [Google Scholar] [CrossRef]
- Hore, A.; Ziou, D. Image quality metrics: PSNR vs. SSIM. In Proceedings of the 2010 20th International Conference On Pattern Recognition, Istanbul, Turkey, 23–26 August 2010; pp. 2366–2369. [Google Scholar]
- Khalel, A. Sewar: A Python Package for Image Quality Assessment Using Different Metrics. GitHub Repository. 2020. Available online: https://github.com/andrewekhalel/sewar (accessed on 20 February 2023).
- De Pittà, M.; Berry, H. Computational Glioscience; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Cheng, C.; Wang, Y.; Xu, L.; Liu, K.; Dang, B.; Lu, Y.; Yan, X.; Huang, R.; Yang, Y. Artificial astrocyte memristor with recoverable linearity for neuromorphic computing. Adv. Electron. Mater. 2022, 8, 2100669. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasenko, S.V.; Kazantsev, V.B. Information Encoding in Bursting Spiking Neural Network Modulated by Astrocytes. Entropy 2023, 25, 745. https://doi.org/10.3390/e25050745
Stasenko SV, Kazantsev VB. Information Encoding in Bursting Spiking Neural Network Modulated by Astrocytes. Entropy. 2023; 25(5):745. https://doi.org/10.3390/e25050745
Chicago/Turabian StyleStasenko, Sergey V., and Victor B. Kazantsev. 2023. "Information Encoding in Bursting Spiking Neural Network Modulated by Astrocytes" Entropy 25, no. 5: 745. https://doi.org/10.3390/e25050745